









Abstract
RNA interference (RNAi) mediates antiviral defense in many eukaryotes. Caenorhabditis elegans mutants that disable RNAi are more sensitive to viral infection. Many mutants that enhance RNAi have also been identified; these mutations may reveal genes that are normally down-regulated in antiviral defense. About one-third of the score of mutants that enhance RNAi are in synMuv B genes, identified 30 years ago in unrelated screens for increased growth factor signaling. Many synMuv B genes encode dREAM complex chromatin-regulatory proteins found in nearly all animals and plants. We show that mRNAs which are highly induced in synMuv B mutants are congruent with those induced by Orsay RNA virus infection, suggesting that the enhanced RNAi of synMuv B mutants may also be triggered by down-regulation of synMuvB gene activity in an Orsay virus infection of wild type. The multivulval (Muv) phenotype of synMuv B mutants requires the presence of a second nematode-specific synMuv A gene mutation, but the enhanced RNAi of synMuv B mutants does not require a second synMuv A mutation. To test if Orsay viral infection down-regulates synMuv B gene activity, we infected a single synMuv A mutant with Orsay virus and found that a Muv phenotype could be induced. Thus, decreased synMuv B gene activity is part of the normal C. elegans viral defense response. In support of the model that decreased syn- Muv B gene activity enhances antiviral response, we found that synMuv B mutants have 50 to 100× lower viral RNA levels during an Orsay virus infection than wild type. Thus down-regulation of synMuv B activity to enhance RNAi is a key component in the defense response to viral infection. Small RNA deep sequencing analysis of dREAM complex mutants revealed siRNA profiles indicative of such a response. Thus, the pan-eukaryotic synMuv B genes constitute an element in C. elegans antiviral defense which is conserved across many eukaryotes where it also may act in viral defense. The enhanced RNAi and conservation of the dREAM complex mutants suggests new therapeutic avenues to boost antiviral defenses.
Citation: Seetharaman A, Galagali H, Linarte E, Liu MHX, Cohen JD, Chetal K, et al. (2025) Decreased SynMuv B gene activity in response to viral infection leads to activation of the antiviral RNAi pathway in C. elegans. PLoS Biol 23(1):
e3002748.
https://doi.org/10.1371/journal.pbio.3002748
Academic Editor: Andrew Mehle, University of Wisconsin-Madison, UNITED STATES OF AMERICA
Received: July 3, 2024; Accepted: December 17, 2024; Published: January 29, 2025
Copyright: © 2025 Seetharaman et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Data Availability: All relevant data are included within the paper and its Supporting Information files. The raw data underlying the figures have been compiled into an Excel file and uploaded as S1 Raw Data. Raw microscopy images have been deposited in Zenodo and are publicly accessible at DOI: 10.5281/zenodo.14289232. The small RNA sequencing reads and associated metadata generated during this study have been deposited in the GEO database under the project accession number GSE283983. Individual sample accession numbers are as follows: GSM8675299 to GSM8675314. Details regarding the data and analyses can be found in the Results and Materials and Methods sections of the manuscript.
Funding: This work was supported by NIH GM044619 to G.R. and NIH R35GM119775 to T.A.M. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing interests: The authors have declared that no competing interests exist.
Abbreviations:
dsRNA,
double-stranded RNA; FDR,
false discovery rate; RdRP,
RNA-dependent RNA polymerase; RNAi,
RNA interference
Introduction
RNA interference (RNAi) is a key eukaryotic defense against viruses. In the nematode Caenorhabditis elegans, the RNAi pathway is multifaceted, with an expanded set of 25 Argonaute proteins of many classes compared to the 8 Argonautes from 2 classes of many animals, including humans. Argonaute proteins bind to small RNAs, including siRNAs or miRNAs of 22 to 26 nucleotides (nt) which guide those Argonaute/siRNA complexes to target mRNAs with complementarity to the siRNAs to induce their degradation or other forms of regulation such as regulation of translation by most miRNAs. In plants, fungi, nematodes, bivalves, and some insects, but not in most animal species, there is a second-stage amplification of siRNAs by the RNA-dependent RNA polymerase (RdRP) proteins. RdRPs work in concert with Dicer, which initially generates 22 to 26 nt primary siRNAs with a 5′ OH which are extended by RdRPs to produce a far more abundant pool of 22 nt secondary siRNAs with 5′ phosphates. This amplification process relies on RdRP using mRNA templates to extend primary siRNAs, often targeting transposons, integrated viruses, and newly invading viral elements. Mutations in either Argonaute or RdRP genes can disable this RNAi defense [1]. This two-stage RNAi system of C. elegans is the probable ancestral state of eukaryotic RNAi, with the as yet unexplained deletion of the RdRP genes from most animals (such as humans) and many fungi, but its retention in all plants and a rather peculiar set of animals that includes the nematodes.
Because of this 2 stage siRNA production pathway, RNAi in C. elegans is remarkably potent, and can even be triggered simply by ingestion from the intestine of double-stranded RNA (dsRNA) produced in an Escherichia coli strain engineered using T7 RNA polymerase promoters flanking a cloning site to produce an approximately 1 kb long dsRNA derived from a C. elegans genomic or cDNA clone [2]. Extensive genetic screens for RNAi-defective (Rde) C. elegans mutants revealed loss of function mutations in dozens of genes that disable RNAi, including in the Argonaute protein gene rde-1 (RNAi defective mutant 1) and a collection of mutator genes defined before the discovery of RNAi as mutations that activate transposition of C. elegans transposons which are retroviral elements, genes with a viral history [3,4]. Given that even ingestion of dsRNA can trigger potent RNAi, it was a surprise that soon after the discovery of RNAi mutations emerged from genetic screens that actually enhance the capacity of C. elegans to silence mRNAs: mutations in a distinct set of 20 genes emerged from genetic screens for enhanced response to dsRNAs that actually enhance the already potent RNAi of C. elegans [5–13]. A significant fraction of the genes that enhance RNAi (Eri) are members of the synMuv B class genes that were previously identified in totally unrelated genetic screens in the Horvitz and Han labs for increased EGF/Ras signaling in patterning the C. elegans cell lineage during development. Among the initial collection of multivulva or Muv mutants from 40 years ago was a set of mutants that carried 2 loss of function mutations, in lin-8 and lin-9, or lin-15A and lin-15B, and required loss-of-function alleles of both genes in order to show the Muv phenotype of increased EGF/ras signaling in the ventral hypodermal lineages [14–16]. Further genetic screens for new mutations that were synthetic Muv with either the lin-8 (a synMuv A mutant) or lin-9 (a synMuv B mutant) mutants, but not as single new mutations, revealed a total of about a dozen synMuv B genes that interact with this growth factor signaling pathway. These genes were classified as either a synMuv A (for example, lin-8 and lin-56) or a synMuv B mutation; only a combination of synMuv A plus a synMuv B mutant caused a multivulval phenotype [14].
Molecular analyses of many synMuv B genes mostly by the Horvitz lab showed that many of them encode homologues of the dREAM complex, which is conserved across animals and plants. These include LIN-35, an orthologue of the human tumor suppressor gene retinoblastoma (Rb), and the Rb complex components LIN-53/RbAp48, LIN-37/MIP40, LIN-54/MIP120, DPL-1/DP, and LIN-9/MIP130 [15–21]. Molecular analyses of synMuv A genes showed that lin-8, lin-56, and lin-15A encode widely expressed nuclear proteins with no obvious homologues outside of nematodes [17,18]; the synMuv A genes either evolve very quickly or have evolved in nematodes or lost in other taxons.
The Muv phenotype of the synMuv A; synMuv B double mutants is caused by abnormally high expression of the LIN-3 growth factor ligand in the hypodermal cells that signal to the adjacent vulval cells. Loss of function mutations in any synMuv B gene in combination with any synMuv A mutant cause excessive LIN-3 EGF growth factor signaling from the hypodermal cells to induce a Muv phenotype in the adjacent vulval cell lineages via the EGF receptor and downstream kinase signaling in those cell lineages [15,18,22,23]. Tissue-specific analysis of synMuv A and synMuv B gene activities strongly implicated their function in the hypodermis to regulate the production of the ligand LIN-3 and for this EGF growth factor signaling pathway in the adjacent vulval cell lineages [24,25]. Tissue-specific expression of the lin-35 synMuv B gene only in the hyp7 hypodermis is sufficient to rescue the Muv phenotype of a lin-8; lin-35 double mutant, whereas expression of lin-35 in the ventral hypodermal cells where the LIN-3 EGF signals are received does not rescue the Muv phenotype [22]. In situ mRNA hybridization of lin-3 shows a dramatic increase in lin-3 mRNA in the hypodermis only in a synMuv A; synMuv B double mutant, but not in either single mutant [15].
SynMuv B dREAM complex components constitute about 1/3 of the genes that have emerged from screens for enhanced RNAi [5,9,10,12]. The previously characterized roles of the synMuv B dREAM complex in cell cycle and transcriptional regulation/chromatin does not obviously intersect with production or response to siRNAs produced during RNAi; the molecular basis of how the DNA-binding proteins and associated proteins of the synMuv B pathway actually intersect with the Argonaute proteins and RdRPs that are central to RNAi has not yet emerged. Further, while the analysis of the synMuv A and synMuv B mutants in EGF signaling from the hypodermis to the ventral precursor cells has been extensive and beautiful, how it connects to the Eri response of the synMuv B mutants is not at all obvious from this molecular genetic analysis of LIN-3 signaling. There are some differences between the hypodermal to vulval cell signaling by lin-3 and the enhanced RNAi of synMuv B mutants: while both a synMuv A and a synMuv B class mutation are needed to increase LIN-3 signaling from hyp7 for vulval induction, only a synMuv B mutation is required to enhance RNAi [7], and there is no entanglement of the synMuv A mutations in either the enhanced or defective RNAi phenotypes.
The pleiotropies of C. elegans dREAM complex mutations suggest a function in the intestine and other non-germline tissues on the production of P-granules, the phase-separated granules initially studied by Priess [26], and by Brangwynne and Hyman, which mediate RNAi, for example Mutator-granules and Z-granules [27,28]. Loss of function mutations in many different C. elegans synMuv B genes cause misexpression of these normally germline-specific P-granules in somatic tissues, most dramatically in the polyploid intestine and hypodermis [7,29,30]. Because RNAi is normally far more active in the germline [31], the somatic activation of the germline P-granule production is consistent with the increased response to dsRNAs that target somatic cells in the synMuv B mutants.
The P-granule misexpression in the somatic intestine is also salient to RNAi because many RNAi components are localized to these perinuclear, phase-separated organelles, and many proteins and RNAs that mediate RNAi localize to P-granules and to the adjacent Mutator granules [32] and Z granules [33], which are also central to RNAi. The gene expression changes in synMuv B mutants supports the model that they regulate particular genomic client loci that mediate RNAi. For example, in L1 stage lin-35(n745)/retinoblastoma mutant animals with just 2 quiescent germline precursor cells (Z2 and Z3) that divide much later in development to generate thousands of germline cells, the P granule genes pgl-3 (up 28×) and pgl-1 (up 10×) and the heritable RNAi Argonaute factor hrde-1 (8×) are significantly up-regulated compared to wild-type L1 stage [34]. These same dramatic gene inductions were also observed in multiple other synMuv B class mutants [29]. But this is not a misspecification of intestinal cell fate to germline fate: the animal is viable and the function of the intestine for nutrition and for vitellogenin synthesis and export to the oocyte is intact. This synMuv B mutant intestinal and hypodermal misexpression of P-granules does not depend on a second synMuv A mutation, unlike the increased EGF signaling in the vulval precursor cells [15].
A common theme to the tissues most affected by the synMuv B mutations, the hypodermis and the intestine, is dramatic endoreduplication. The adult C. elegans intestine is normally endoreduplicated from diploid at hatching to 32C, 5-inferred full genome duplications; with one duplication at each larval stage but no chromosome condensation or mitosis [25]. The hyp7 hypoderm where ectopic P granules are also observed is 8C [35]. Thus, 2 endoreduplicated tissues are affected by dREAM complex genes. Local gene amplification by the dREAM complex has also been observed in Drosophila where the dREAM complex binds at the Drosophila chorion genes to control a 50 kb endoreduplication to nearly 100× the gene dosage [36]. This enables the production of the abundant eggshell proteins; mutations in the dREAM complex cause a humpty dumpty phenotype, fragile eggs due to insufficient production of chorion proteins [37–39].
The enhanced RNAi phenotype of the many synMuv B and other Eri mutants, and the siRNA and gene expression changes associated with eri-gene mutations points to an elaborate regulation of RNAi activity; the synMuv B mutations which hyperactivate RNAi may place this regulated pathway in a constitutively ON antiviral state, when it is normally a transient state during an actual viral infection. That RNAi may be highly regulated is not surprising: RNAi has evolved by selection for potent antiviral defense during almost a billion years of eukaryotic infection by viruses. But it is surprising that the dREAM complex is so central to RNAi.
Here, we show that C. elegans synMuv B mutations activate viral defense pathways, leading to gene expression normally associated with viral infection, but in the absence of virus exposure. We show that Orsay RNA virus infection of C. elegans causes highly congruent gene expression responses to those of synMuv B mutants. Most importantly, we show that a viral infection induces a physiological state that down-regulates synMuv B gene activity: like the synthetic multivulval phenotype that is normally only observed when C. elegans carries both a synMuv A and synMuv B mutation, Orsay viral infection of a synMuv A mutant causes a partially penetrant Muv phenotype. Importantly, this physiological state confers resistance to Orsay virus: infected synMuvB mutants support 50 to 100× lower Orsay virus RNA levels compared to wild-type animals. These data together show that the down-regulation of synMuv B gene activity or dREAM function is a key defense response that is triggered during an actual viral infection. Thus, the mammalian-conserved synMuv B pathway highlights new avenues to activate production or response to siRNA and may be targeted with drugs to activate human antiviral responses or responses to RNAi-based pharmaceuticals.
Results
synMuv B null mutants turn on many genes that are up-regulated during viral infection
Enhanced RNAi (Eri) mutants were first identified by their activation of RNAi in neurons, for example, which normally do not recapitulate the phenotypes caused by loss of function mutations in the corresponding genes. For example, RNAi of various synaptic components often cause no phenotype in wild type, even though genetic mutations cause strong movement disorders; for still unknown reasons neurons in wild type are deficient in RNAi [40]. The eri mutants also show enhanced responses to a set of feeding dsRNAs that were discovered over the years which did not recapitulate expected loss of function phenotypes; lin-1, hmr-1, lir-1, cel-1, unc-73, or dpy-13 RNAi do not cause phenotypes in wild type but strong phenotypes in eri mutants [10]. One model for the existence of the eri mutants is that each of them constitutively activates what is normally a highly transient state of increased RNAi, for example, during a viral infection. We explored whether the Eri response of synMuv B mutants might be evidence of their involvement in the orchestration of a coordinated genetic response towards a perceived viral threat, rather than a mere byproduct of gene expression-related abnormalities.
To explore this hypothesis, we analyzed mRNA seq data sets for C. elegans synMuv B lin-35(n745) and lin-15B(n744) null mutants (available through the NCBI GEO database collection), for gene expression signatures that may show similarities to the gene expression responses to infection of C. elegans by the Orsay RNA virus. We analyzed the mRNA seq data sets of developmental-stage matched wild-type N2 and synMuv B lin-35 and lin-15B mutants for the induction of genes that are strongly induced during Orsay viral infection (GEO accession numbers in Materials and methods). About 90 C. elegans genes are 3- to 10-fold up-regulated when animals are infected by the Orsay RNA virus compared to control animals [41]. A highly overlapping set of 50 genes are 4- to 10-fold up-regulated in lin-15B or lin-35 L1 stage animals compared to wild-type L1 animals. For example, 20 of the 50 lin-15B mutant-induced genes and 22 of the 50 lin-35 mutant-induced genes are also induced in wild-type animals upon Orsay virus infection. Many of the most highly virus-induced C. elegans response genes and most highly induced lin-15B or lin-35 mutant response genes are the pals genes [42,43], which are also dramatically up-regulated in both lin-35(n745) and lin-15B(n744) null mutants (Table 1 and S1 Fig). Each of the differentially expressed genes shown in Table 1 and S1 Fig are 5-fold or more induced in the synMuv B mutants compared to wild-type controls in the absence of any viral infection. We list the top 50 genes that are up by at least 5-fold in lin-15B(n744) and lin-35(n745), mutants in S1–S3 Tables. Perhaps indicative of a local chromatin regulatory mechanism, the 39 PALS genes are localized in five, 20 to 50 kb clusters in the C. elegans genome [42–45]. Significantly, a mutation in the pals-22 gene causes increased transgene silencing, a common enhanced RNAi mutant phenotype, including the synMuv B mutants [42–45]. The clustering of these genes may be related to their potential co-regulation or co-amplification during a response to a viral infection.

Table 1. Differentially expressed C. elegans genes upon Orsay virus infection are up-regulated in lin-35(n745) and lin-15B(n744) synMuv B mutants under non-infection conditions.
Listed here are 17 genes previously identified as top statistically significant but functionally uncharacterized in Orsay-infected C. elegans [43], and their level of expression within and their level of expression within lin-35(n745) or lin-15b(n744) synMuv B mutants under non-infection conditions within the mRNA seq data sets GSM4697089- lin-35[JA1507(n745) rep1/L1, GSM4697090- lin-35[JA1507(n745) rep1/L1], GSM1534086- lin-35[JA1507(n745) rep1 L3 and GSM1534087- lin-35[JA1507(n745) rep1 L3], and GSM4697102- lin-15b(n744)- rep1/L1, GSM4697105- lin-15b(n744)- rep2/L1], that were obtained from the NCBI Geo collection. The log2 gene expression Fold changes shown for N2 animals upon Orsay virus infection were obtained from Chen and colleagues [40].
A pals-5 promoter fusion to GFP has been developed as a reporter of viral infection response by the Wang and Troemel labs after their RNA seq of viral and microsporidia infection showed that pals- genes are highly induced [42]. The strong induction of pals- genes in the lin-15B or lin-35 synMuv B mutants may report the aberrant induction of antiviral defense in the absence of a real viral infection in these mutants. For example, a pals-5p::GFP fusion gene is not expressed in wild-type C. elegans not infected with Orsay virus, but is strongly activated in the intestine of a wild-type animal carrying the pals-5p::GFP after Orsay virus ingestion, the mode of C. elegans Orsay viral infection [42]. To explore this viral response in the synMuv B mutants, we used RNAi or mutations to disrupt particular synMuv B genes in a strain carrying pals-5p::GFP. RNAi of the synMuv B genes lin-9 or lin-13 strongly activated pals-5p::GFP (Fig 1A and 1B). We also crossed strong loss-of-function mutations of lin-35(n745), lin-52(n771), and lin-9(n112), into a strain carrying pals-5p::GFP and found that all the 3 synMuv B homozygous mutants robustly activate pals-5p::GFP (Fig 1C). We further tested by RNAi the synMuv A genes lin-8 or lin-56 as well as several lido (LIN-8 domain containing) genes, paralogues of the synMuv A gene lin-8 [46], but these gene inactivations of synMuv A genes or their paralogues did not induce pals-5p::GFP (Fig 1B). These findings support the hypothesis that the C. elegans synMuv B mutants may place the animal in a physiological state that is normally induced by a viral infection, triggering the activation of their antiviral response, for example, induction of specific viral defense genes and activation of RNAi.

Fig 1. A pals-5p::GFP fusion gene is induced in synMuv B mutants in the absence of viral infection.
(A) Fluorescent micrographs showing pals-5p::GFP expression in animals raised on E. coli expressing dsRNA of empty vector control L4440 or dsRNA targeting lin-9 or lin-13 synMuv B genes. Scale bar is indicated. (B) Quantification of the percentage of animals showing pals-5p::GFP expression when raised on E. coli expressing dsRNA against a panel of different synMuv B eri-genes including lin-9, lin-13, and non-Eri synMuv A genes lin-8, lin-56, and a panel of lido (for lin-8 domain) genes (C). Quantification of the percentage of animals showing pals-5p::GFP expression in lin-35(n745); pals-5p::GFP, lin-9(n112); pals-5p::GFP and lin-52(n771); pals-5p::GFP synMuv B mutants and percentage of lin-35(n745); pals-5p::GFP animals showing pals-5p::GFP expression when raised on E. coli expressing dsRNA against the synMuv Eri suppressor genes isw-1 and mes-4 and other key genes that encode components central to RNAi such as rde-1, rde-4, and mut-16 (D). Percentage of animals showing pals-5p::GFP expression when raised on E. coli expressing dsRNA against the underlined panel of genes in the jyIs8[pals-5p::GFP] animals. Non-underlined genes are genetic doubles that were generated in the pals-5p::GFP background with strong loss-of-function mutations of the genes shown. Statistical comparisons were made between samples marked with asterisks and the sample denoted by a solid pink dot. **** indicates p p S1 Raw Data. Raw microscopy images corresponding to panel (A) have been deposited in Zenodo and are accessible at DOI: 10.5281/zenodo.14289232.
Also suggesting that the pals gene family act in a pathway of anti-viral defense, multiple pals-22 null mutations were identified in a genetic screen for silencing of multicopy transgenes, which are recognized by the RNAi pathway as foreign genetic elements akin to viruses and silenced by the pathway [45]. Transgenes are recognized as foreign, most especially in Eri mutants such as the synMuv B and other Eri mutants (a concept that is further assessed in later experiments discussed in this study) [10]. The pals-22 allele that emerged from a screen for transgene silencing also showed enhanced RNAi with the dpy-13 and cel-1 feeding RNAi that are standard tests for enhanced RNAi [45]; thus, the pals-22 loss of function mutant is an Eri mutant. The induction of the pals genes by viral infection and by the synMuv B mutations in lin-15B or lin-35, and the observed enhanced RNAi of lin-15B or lin-35 null mutants is consistent with association between pals- genes and antiviral defense.
Gene inactivations that suppress the Muv phenotype of synMuv B; synMuv A double mutants also suppress pals-5p::GFP induction by synMuv B loss of function mutations
The Han group discovered 32 gene inactivations that can suppress the Multivulvae (Muv) phenotype of synMuv A; synMuv B double mutants [14]. Seven of those Muv suppressor gene inactivations, mes-4, isw-1, gfl-1, mrg-1, ZK1127.3, M03C11.3, and C3410.8 were nearly 100% suppressors of the Muv phenotype by RNAi. A subset of these synMuv suppressors, also suppress the Eri phenotype of synMuv B mutants [14]. Two of the more potent synMuv B Eri suppressors include isw-1, which encodes a member of the ISW1 complex and mes-4, which encodes a Set domain containing protein, fitting with the chromatin annotation of the dREAM complex [14]. We tested whether the induction of pals-5p::GFP in synMuv B mutants is functionally linked to their Eri phenotype. We reasoned that suppressing the Muv or the Eri phenotype of synMuv B mutants could also cause a decrease in the induction of the pals-5p::GFP reporter. To test this, we targeted isw-1 and mes-4 Muv suppressors by RNAi in the lin-35(n745); pals-5p::GFP strain. As controls, we also targeted other core components of the RNAi machinery such as the Argonaute gene rde-1, the RNA helicase gene rde-4 and the mutator protein gene mut-16 and determined the percentage of GFP-positive animals in their F1 progeny. We found that knocking down isw-1, mes-4, rde-1, rde-4, and mut-16 by RNAi partially but not completely suppressed the percentage of GFP-positive animals in the lin-35(n745); pals-5p::GFP animals (Fig 1C). We also showed that inactivation of isw-1, mes-4, rde-1, rde-4, or mut-16 by RNAi in an otherwise wild-type background does not activate pals-5p::GFP (Fig 1D).
We also characterized the induction of pals-5p::GFP in enhanced RNAi mutants that are not synMuv B genes and found that only synMuv B mutations or synMuv B gene RNAi induce pals-5p::GFP. For example, inactivation by RNAi or by mutations of the eri genes eri-6, eri-6/7, eri-1, and rrf-3 does not activate pals-5::GFP (see Fig 1C and 1D). This suggests that the Eri phenotype induced by synMuv B gene mutations may enhance RNAi via a pathway that is distinct from the other known Eri mutants. In addition, targeting prg-1, the worm ortholog of PIWI, which silences integrated viruses in Drosophila [47], by RNAi, also did not activate pals-5::GFP expression (see Fig 1D), suggesting that piRNA-mediated defense response is distinct from that of the Eri phenotype caused by synMuv B mutants.
We also infected pals-5p::GFP wild type and lin-35(n745); pals-5p::GFP animals with the Orsay virus and examined their F2 progeny for pals-5p::GFP activation. We examined F2 progeny to allow sufficient time for the viral infection to become well-established within the population. pals-5p::GFP animals infected with the Orsay virus showed a robust activation of pals-5p::GFP, whereas pals-5p::GFP animals not infected with Orsay virus did not induce GFP expression (Fig 2A and 2C). The lin-35(n745); pals-5p::GFP animals show 100% penetrant GFP activation under both infected and uninfected conditions (Fig 2A–2C). Inactivation of isw-1 and mes-4 by RNAi in lin-35(n745); pals-5p::GFP animals under non-Orsay infection conditions significantly suppressed the pals-5p::GFP reporter activation (Fig 2A, 2B, and 2D). However, upon infection by the Orsay virus, we no longer observed this suppression, which suggests that isw-1 and mes-4 function while required for the induction of pals-5p::GFP in lin-35(n745) null mutants under non-infection conditions may be dispensable or insufficient in the context of an actual viral infection.

Fig 2. Gene inactivation by RNAi of the synMuv suppressor genes encoding the chromatin remodeling and histone methylation factors isw-1 and mes-4 significantly suppresses the activation of pals-5p::GFP in the lin-35(n745) mutant under non-infection conditions but does not suppress pals-5p::GFP response to Orsay virus infection.
(A) Fluorescent micrographs of pals-5p::GFP (control) or lin-35(n745); pals-5p::GFP animals not exposed to Orsay virus or after Orsay virus infection. A brightfield image highlights the number of animals that were imaged. Scale bar is indicated. (B–D) Graphs showing a quantification of pals-5p::GFP(control) or lin-35(n745); pals-5p::GFP under RNAi of isw-1 or mes-4, as well as failure of isw-1 or mes-4 RNAi to suppress induction of pals-5p::GFP by Orsay virus infection. **** indicates p S1 Raw Data. Raw microscopy images shown in this figure have been deposited in Zenodo and are accessible at DOI: 10.5281/zenodo.14289232.
lin-15B null mutants exhibit altered intestinal nuclear morphology
C. elegans infection with Orsay virus causes morphological changes in intestinal cells, including cell fusion, elongation of nuclei, and nuclear degeneration [48]. We looked for similar phenotypes using 2 strains that carry transgenes to visualize the intestinal nuclear membrane using the mCherry tagged nuclear pore protein NPP-9 (ges-1p::npp-9::mCherry::BLRP::3xflag or ges-1p::npp-9::mCherry::BLRP::3xFLAG; his72p::birA::GFP). One set of animals were infected with Orsay virus while the control group received no virus, and both groups were observed for 2 generations. The intensity of viral infection was established by the robust expression of pals-5p::GFP in animals carrying that transgene and infected simultaneously (Fig 3C, top panel). The no virus infection control animals had an average of 33.2 ± 2 intestinal nuclei at the adult stage. Orsay virus infected adults exhibited a 5% decrease in the number of intestinal nuclei, (31.5 ± 2.5) (Fig 3A and 3B), weakly supportive of the previously observed nuclear degeneration. We also observed elongation of intestinal nuclei in some Orsay virus-infected animals (Fig 3C, middle panels).

Fig 3. lin-15b(-) animals exhibit altered intestinal nuclear morphology.
(A) Number of mCherry expressing intestinal nuclei in FAS207 adults in wild-type and lin-15b(W485*) mutant backgrounds. (B) Number of mCherry expressing intestinal nuclei in JJ2284 adults in WT and lin-15b(W485*) backgrounds. The square represents the average, and the bars represent the standard deviation. The sample size n has been indicated; p value calculated using ANOVA with Bonferroni correction. n.s: not statistically significant, * p p C) Representative images of pals-5p::GFP (top row), FAS207, and lin-15b(W485*); FAS207 adults. White arrows indicate elongated intestinal nuclei. Scale bar is indicated. The graphs shown in A and B were generated using custom scripts in R studio and the statistical analyses were done using Jupyter Notebook. The numerical data underlying the graphs shown in this figure can be found in the Supporting information data table named S1 Raw Data. The raw microscopy images shown in this figure have been deposited in Zenodo and are accessible at DOI: 10.5281/zenodo.14289232.
To further test the intestinal nuclear response in synMuv B mutants to Orsay virus infection, we constructed a lin-15B(W485*) null mutant allele based on the canonical lin-15B(n744) null allele using CRISPR in the strain harboring the transgene ges-1p::npp-9::mCherry::BLRP::3XFLAG. We observed that under no virus infection conditions, lin-15B(W485*) animals exhibited a 15% decrease in the number of intestinal nuclei, with an average of 28.1 ± 3.5 nuclei (Fig 3A). L1 larval stage wild-type animals normally have 20 intestinal nuclei which increase to 36 nuclei by the L2 stage, as most L1 intestinal cells undergo mitosis or at least nuclear division. Those 36 nuclei then increase their DNA content by undergoing DNA replication without cytokinesis during the next 2 larval stages and into adulthood, ending up with 36 nuclei with 32C genome copy number as roughly assayed by DAPI staining of DNA [35]. Thus, the minimum number of nuclei we could expect to observe if there was no cell division at the first larval stage is 20; so, a decrease from 33 nuclei in wild type and to 28 nuclei in a lin-15B null mutant was substantial.
Strikingly, many of the intestinal nuclei in the lin-15B(W485*) animals were elongated (Fig 3C, lower panels), suggesting this Orsay virus-associated cellular alteration is constitutively present in these lin-15B(-) animals. The decrease in the number of intestinal cells and the altered morphology of intestinal nuclei was also observed in a second independent line of lin-15B(W485*) that was generated in the JJ2284 strain (S8 Fig). Orsay virus infection of the lin-15B(W485*) animals did not cause a further reduction of the number of intestinal nuclei or any change in the intestinal nuclear morphology. This implies that the intestinal nuclear response to viral infection of wild-type C. elegans may be a direct result of down-regulation of lin-15B or other synMuv B genes in Orsay virus-infected animals. Fusion of intestinal cells in Orsay virus-infected animals has been reported previously [48]. Both the observed decrease in the number of nuclei as well as the elongated appearance of the intestinal nuclei could be the result of a fusion of intestinal nuclei. However, we cannot rule out the alternate possibility that the decrease in the number of intestinal nuclei in lin-15B(-) animals and Orsay virus-infected animals could be due to a lineage defect or defect in karyokinesis during development.
Orsay virus infection down-regulates synMuv B gene activity
The C. elegans Delta-like ligand for the GLP-1 and LIN-12 Notch receptors is encoded by lag-2. lag-2 is normally expressed only in the distal tip cells of the gonad and a subset of the vulval precursor cells [49]. But mutations in several synMuv B genes cause nearly 100% penetrant ectopic lag-2::GFP expression in intestinal cells, which can be suppressed by gene inactivation of synMuv suppressor genes such as isw-1 [14]. If a C. elegans viral infection normally triggers the down-regulation of synMuv B genes, perhaps as a natural defense mechanism, we hypothesized that this may cause misexpression of lag-2 within the intestinal cells as is observed in non-virally infected synMuv B mutants [14]. We infected wild-type animals carrying the lag-2p::GFP fusion gene with the Orsay virus and scored their F2 progeny for lag-2p::GFP misexpression in the intestine during the L4/adult stages. Upon infection of wild-type animals with the Orsay virus, lag-2p::GFP was misexpressed in the intestinal cells with a 100% penetrance, similar to synMuv B mutants that are not infected with Orsay virus (Fig 4A). This virus-induced intestinal misexpression of lag-2p::GFP was not suppressed by inactivation of synMuv suppressor genes such as isw-1, in contrast to the lag-2p::GFP induction in the synMuv B mutants (S2A and S2B Fig). These data show that infection with Orsay virus recapitulates certain synMuv B loss of function phenotypes, for example, the induction of lag-2p::GFP in the intestine.

Fig 4. Orsay virus infection causes misexpression of lag-2p::GFP in somatic tissues.
(A) Shown are fluorescent micrographs depicting lag-2p::GFP expression in worms under no virus and under Orsay virus infection. The position of the distal tip cells of the germline are indicated by yellow arrows and the panels to the right of the WT/uninfected and WT/infected worms are panels of the region that is boxed on the left at a higher magnification. New ectopic expression of GFP seen within intestinal cells in the virus-infected animals is labeled with red arrows. Ectopic expression of GFP within intestinal cells of lin-35(n745) and lin-9(n112) mutants is shown as additional positive controls and the ectopic expression of GFP in the intestine is labeled with red arrows. The position of the central vulva is labeled with a white arrow. Scale bar is indicated. (B) Shown here is a quantification of the percentage of Rol phenotype of mgIs30 animals that bear an integrated rol-6 transgene, under no virus and infection with the Orsay virus. The suppression of the rol-6 transgene in a lin-35(n745) null mutant is shown to provide a quantitative estimate of the intensity of the enhanced RNAi phenotype that is typical of most Eri mutants. Statistical comparisons were made between samples marked with asterisks and the sample denoted by a solid pink dot. **** indicates p 10.5281/zenodo.14289232.
An integrated multicopy transgene (mgIs30) carrying a tandemly duplicated collagen mutant allele rol-6(su1006), an R71C substitution mutation of rol-6 that causes the Rolling phenotype due to collagen assembly defects [50], causes a nearly 100% rolling (Rol) phenotype in wild type. But in nearly all Eri mutants (eri-1, rrf-3, eri-6/7, lin-35, lin-15B, and other synMuv B mutants), this transgene, like most multicopy transgenes, is recognized as foreign and silenced so that the animals no longer roll [11]. We reasoned that if down-regulating synMuv B genes represent a natural part of the antiviral defense, then a viral infection itself may induce an Eri response, which may silence multicopy transgenes such as the rol-6 transgene. We infected a wild-type strain carrying the mgIs30 multicopy rol-6 transgene with the Orsay virus and scored the percentage of rolling animals in the F2 generation. Consistent with this hypothesis, Orsay viral infection partially silenced the Rol phenotype of mgIs30 (Fig 4B), though to a weaker extent than what is observed in a synMuv B Eri mutant such as a lin-35 null mutant, for example, where the silencing of the rol-6 transgene is nearly 100% (Fig 4B). This difference in the level of transgene silencing may be due to the extent of inhibition of synMuv B gene activity, for example, due to variations in viral titer or the synMuv B pathway response to a viral infection.
Orsay virus infection of a synMuv A null mutant induces a Muv phenotype
Given that lag-2p::GFP is strongly misexpressed with a 100% penetrance within intestinal cells in wild-type C. elegans infected with Orsay virus (which is strikingly similar to the lag-2p::GFP intestinal expression observed in synMuv B mutants), and the observation that Orsay virus infection in wild-type animals enhances RNA interference, we hypothesized that down-regulation of some or all synMuv B genes may represent a normal step in viral defense. The simplest prediction of this hypothesis is that infecting a synMuv A mutant with the Orsay virus might down-regulate synMuv B activity as part of the normal response to a viral infection, so that, like a synMuv A; synMuv B double mutant, viral infection of a synMuv A mutant may cause a Muv phenotype. Only when both synMuv A and synMuv B genes are mutant does the multiple vulvae or Muv phenotype occur in non-virally infected animals. If a viral infection in animals lowers synMuv B gene activity as a defense mechanism, we predicted that Orsay virus infection of a synMuv A null mutant, just the single synMuv A mutation, might be sufficient to cause a Muv phenotype in some fraction of the population.
To test this, we infected the synMuv A lin-8(n2731) null mutant with the Orsay virus and examined F2 progeny for the appearance of the Muv phenotype. Strikingly, in 3 independent trials, we found that 10% of the F2 progeny were indeed Muv (Fig 5A and 5B), whereas uninfected lin-8 mutant control animals did not produce any Muv progeny. The appearance of a Muv phenotype has never been reported in wild-type C. elegans under Orsay virus infection conditions in any previous study nor in our own observations (Fig 5B).

Fig 5. Orsay virus infection of the lin-8(n2731) synMuv A mutant induces a Muv phenotype indicative of decreased synMuv B gene activity, and 2 different synMuv B mutants produce 50–100× less Orsay virus RNA.
(A) Brightfield micrographs showing uninfected vs. virus-infected lin-8(n2731) mutant animals. The central vulva that is normally induced in wild type is indicated by a black arrow. The inset image in the panel showing the virus-infected animal is a magnified region showing ectopic vulval inductions. The ectopic vulval inductions are indicated by red arrows. (B) Quantification of the percentage of Muv animals seen in lin-8(n2731) mutants under uninfected compared to Orsay virus infection. The Muv quantification was performed on the F2 generation of the virus-infected animals. Scale bar is indicated. (C) RT-qPCR for Orsay RNA1 in uninfected wild type N2, infected wild type N2, infected eri-1(mg366), infected mut-16(pk710), infected lin-35(n745), and infected lin-15b(n744) animals. The infected lin-35(n745) and lin-15b(n744) animals produce 50× and 100× less Orsay viral RNA, respectively, after Orsay infection. The gray circles indicate 2 technical replicates from 2 biological replicates. The black square represents the average of the 4 values and the error bars represent the standard deviation. Two-tailed unpaired Student’s t test p-values **p p 10.5281/zenodo.14289232.
To rule out Orsay induction of a synMuv B mutation, we followed the progeny of these Orsay-infected synMuv A mutant animals. We picked single progeny from 8 gravid lin-8(n2731) adult animals that were Muv after an Orsay virus infection, bleach treated them to remove any residual viral particles that may be present, and then allowed their eggs to hatch and examined their progeny at the adult stage to discern if the Muv phenotype persisted. We found that 100% of their progeny had a non-Muv vulval phenotype. Thus, the Orsay virus infection did not induce a mutation in a synMuv B gene in lin-8(n2731) null animals to give rise to a Muv phenotype.
Taken together, these findings suggest that down-regulation of one or more synMuv B genes is a normal step in the antiviral defense response, and that viral infection induces an enhanced RNAi physiological state. The 10% penetrance of the Muv phenotype suggests that the down-regulation of the synMuv B gene activity is just at the threshold for synergizing with the synMuv A mutations to confer a Muv phenotype (see Discussion).
Down-regulation of synMuv B gene activity increases viral resistance
In response to Orsay virus infection, synMuv B gene activity is down-regulated, raising the question of whether this reflects an adaptive antiviral mechanism or a pathogenic consequence of infection. To investigate this further, we infected wild-type or the synmuv B mutants lin-35(n745) or lin-15b(n744) with Orsay virus filtrate at the L1 stage and measured levels of Orsay RNA1 in adults by RT-qPCR, as a proxy for infection levels. If synMuv B gene down-regulation is part of the antiviral response, we predicted that synMuv B mutant animals would display lower infection levels as measured by viral RNA levels. Conversely, if the down-regulation is a pathogenic consequence, mutant animals would exhibit infection levels equal to or greater than those of wild type. lin-35 mutant animals had 50-fold lower and lin-15b mutant animals had 130-fold lower Orsay RNA1 than wild-type animals (Fig 5C). These results suggest that synMuv B mutants are more primed for viral defense, with the reduced synMuv B activity conferring an enhanced antiviral response, perhaps evidence that the animal is in an antiviral physiological state of low synMuv B gene activity.
Given that small RNA pathways are central to the antiviral response in C. elegans [48,51,52], we further analyzed Orsay viral RNA levels in eri-1(mg366), an RNAi-enhanced mutant [40], and mut-16(pk710) [3,30], an RNAi-deficient strain. As anticipated, mut-16 mutants displayed a 10-fold increase in Orsay viral RNA levels, underscoring the essential role of small RNAs in viral defense. Interestingly, eri-1 mutant animals showed only a 2.5-fold reduction in Orsay viral RNA compared to wild type (Fig 5C). This suggests that loss of synMuv B activity results in a far more robust antiviral response than the loss of eri-1. While synMuv B mutants also exhibit enhanced RNAi phenotypes [7], the specific small RNA populations or tissues involved in this stronger antiviral response remain to be determined.
The siRNA landscape of synMuv B Eri mutants
Abundant endogenous siRNAs derived from thousands of genes are produced by wild-type C. elegans that are not infected with viruses or exposed to specific dsRNAs that trigger additional exogenous RNAi of one targeted gene. These natural siRNAs regulate the mRNA levels and chromatin structures of a huge number of genes in wild-type C. elegans; nearly all of these siRNAs are not produced in the RNAi-defective mut-2 or mut-16 mutants, and stunningly, these missing siRNAs from thousands of genes cause almost no phenotypes except for activation of normally dormant transposons and a failure to respond to injected or ingested dsRNAs [3,4,53,54]. In contrast, enhanced RNAi mutants disrupt the production of a far smaller subset of endogenous siRNAs. C. elegans siRNAs are classified by their length and 5′ nucleotide, which generally predict which of the nearly 2 dozen Argonaute proteins present them to specific mRNA and other target RNAs [55]. The primary 26G siRNAs (26 nt in length with G at the 5′ end), associate with the Argonaute proteins ERGO-1 and ALG-3/4. The targets of ERGO-1 26G siRNAs include about 100 protein coding genes, pseudogenes, and long noncoding RNAs, many of which are integrated retrotransposons [54]. The ergo-1 mutants are strongly enhanced for RNAi, suggesting that either silencing these retrotransposons uses much of the RNAi bandwidth of wild-type C. elegans, or that the viral activation that results from failure to silence those retrotransposons with ERGO-1 siRNAs when ergo-1 is defective, in turn enables a viral infection from those normally silenced retroviruses, to in turn induce an antiviral response, or increased RNAi, a sort of C. elegans inflammatory response. Targets of the ALG-3/4 26G siRNAs include spermatogenic genes [56–59]; these 26G siRNAs and the Argonaute that mediate their recognition of mRNAs in sperm have not been implicated in normal RNAi or enhanced RNAi, and the 100× to 1,000× more abundant 22G siRNAs (22 nt in length with G at the 5′ end), are loaded into the many WAGO Argonautes, which are greatly expanded in the nematodes compared to other animals, and CSR-1. A large fraction of these 22G siRNAs is produced in the germline to mediate the massive level of gene silencing in the germline and to ensure that silenced genes from the parent continue to be silenced in progeny. CSR-1-associated 22G siRNAs are also expressed in the germline and modulate expression of germline genes as well as genes that mediate chromosome segregation at meiosis and mitosis [60,61]. The WAGO-associated 22G siRNAs target protein coding genes, pseudogenes, and transposons [27]. Almost all 22G siRNAs are depleted in the mut-16 RNAi defective mutant, but most of these very abundant siRNAs are produced normally in the enhanced RNAi mutants eri-6/7, ergo-1, rrf-3, and eri-1 [10]. But the eri-6/7 and ergo-1 Eri mutants exhibit nearly complete depletion of the 26G siRNAs and a subset of secondary WAGO 22G siRNAs that are produced from these particular primary 26G siRNAs [12,58]. Because SynMuv B mutants exhibit a very strong Eri phenotype that is comparable to that of other well-studied Eri mutants, we investigated the landscape of small RNAs that are generated in these mutants.
Given that many of the key synMuv B mutant phenotypes are observed in polyploid somatic cells (the 8C hypoderm for LIN-3 activation and the 32C intestine for ectopic P-granule formation), we first carried out deep sequencing characterization of all siRNAs in synMuv B double mutants that produce little to no germline: glp-4(bn2), lin-35(n745); glp-4(bn2) and lin-15B(n744); glp-4(bn2) grown at the non-permissive glp-4 temperature of 25°C (Figs 6, S9, and S10). The glp-4(bn2) mutation causes a temperature sensitive genetic ablation of the C. elegans germline when animals are grown at 25°C. The complete list of the significantly up-regulated and down-regulated by a factor of 5-fold siRNAs are presented in the S1 Data and GEO accession numbers for these data sets can be found in the methods section.

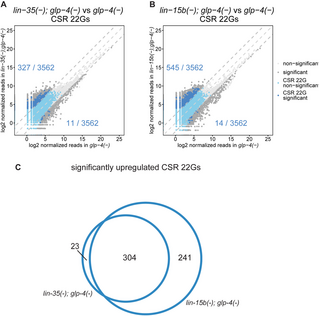
Fig 6. A particular class of CSR 22Gs are up-regulated in synMuv B mutants.
(A, B) Comparison of normalized reads of CSR 22Gs expressed in lin-35(n745); glp-4(bn2) (A) and lin-15b(n744); glp-4(bn2) (B) with glp-4(bn2). Dark gray data points represent small RNAs that are differentially expressed by 5-fold and adjusted p-value C) Venn diagram representing overlap between CSR 22Gs up-regulated in synMuv B mutants in (A) and (B). The numerical data underlying the graphs shown in this figure can be found in the Supporting information data table named S1 Raw Data.
We observed greater than a 5-fold increase in about 10% of the detected CSR-associated 22G siRNAs in both lin-35(n745); glp-4(bn2) and lin-15B(n744); glp-4(bn2) mutants compared to the glp-4(bn2) control animals (Fig 6A and 6B). There was significant overlap between the genes targeted by the up-regulated CSR-22Gs in both lin-35(n745); glp-4(bn2) and lin-15B(n744); glp-4(bn2) mutants, suggesting that a common pathway is misregulated in these distinct synMuv B mutants (Fig 6C). SynMuv B mutants are also known to misexpress many genes that are normally germline-restricted in somatic tissues such as the intestine [7,29]. Because the animals that we analyzed have no germline, the significant increase in CSR-22G siRNAs that are observed in these synmuvB mutants may reflect the somatic misexpression of these germline genes, perhaps in the intestine. We also observed loss of about half of the detected ERGO-26G siRNA in both lin-35(n745); glp-4(bn2) and lin-15B(n744); glp-4(bn2) (S9C and S10C Figs). This partial loss of ERGO-26G siRNAs in synmuvB mutants is less severe than the complete depletion of ERGO 26G siRNAs in eri-1, eri-6, eri-7, and ergo-1 loss of function mutants [12,58]. It is possible that the synMuv B mutants affect 26G siRNA production in the intestine, for example, whereas other eri- mutants may affect more cell types.
WAGO-22G siRNAs were differentially expressed in both lin-35(n745); glp-4(bn2) and lin-15B(n744); glp-4(bn2), with some showing increased and others showing decreased expression (S9G and S10G Figs). We also observed a decrease in the expression of a few microRNAs and piRNAs in both lin-35(n745); glp-4(bn2) and lin-15B(n744); glp-4(bn2) (S9K, S9L, S10K, and S10L Figs). An exciting possibility is that these particular miRNAs, for example, mir-8191 may be located, in locally amplified regions that depend on the dREAM complex, as has been observed for the Drosophila chorion genes [36–39]. Finally, we found very little change in the abundance of ALG class 26G siRNAs (S9A and S10A Figs).
Additionally, we carried out in parallel, the deep sequencing of small RNAs in lin-15B(n744), lin-35(n745), lin-9(n112), and lin-52(n771) synMuv B mutants with intact germlines (for GEO accession numbers for these data sets, see Methods section). We did not detect any striking differential expression of any of the classes of small RNAs, possibly because the abundance of small RNAs derived from the germline may mask the changes in siRNA expression that may occur in the soma (S11–S14 Figs).
Molecular epistasis analysis of the enhanced RNAi in synMuv B mutants
The 26G siRNAs in animals are synthesized via the action of the RdRPs RRF-1,2,3 and EGO-1, which produce longer dsRNAs that are further processed into smaller 26G-RNAs through DICER and PIR-1/phosphatase [57–59,62,63]. Our deep sequencing analysis of the synMuv B mutants is consistent with the decline in 26G siRNAs in most Eri mutants, such as eri-1(mg366) [58] or eri-6/7 and ergo-1 [10,12,64]. To further explore the 26G axis of siRNAs, we mined mRNAseq data sets (from the NCBI Geo-collection) of lin-35(n745) and lin-15B(n744) mutants, to see whether transcripts of any genes that are known to cause enhanced RNAi (such as eri- genes, rrf-3 etc), when mutant were down-regulated in synMuv B mutants. However, we did not observe lower mRNA levels for any of the known genes that mutate to an enhanced RNAi phenotype in the lin-15B and lin-35 mRNA-seq data sets (see S2–S4 Data files). This suggests that the decrease in 26G siRNAs in the synMuvB mutants is not due to the down-regulation of other known eri- genes. 26G-siRNAs are bound by ERGO-1 Argonaute which then targets those siRNAs to particular target mRNAs, approximately 100 recently acquired retroviral elements [10–12]. To test whether synMuv B mutations affect ERGO-1 protein levels or localization to in turn affect 26G siRNA levels, we observed the ERGO-1 protein using a GFP-tagged full-length ERGO-1 protein (tor147[GFP::3XFLAG::ergo-1, the longest predicted isoform [65]. We used RNAi to inactivate synMuv B genes or to inactivate genes that encode suppressors of synMuv B genes, and then observed the localization and expression of GFP::3XFLAG::ERGO-1 to determine if down-regulating the activity of either synMuv B genes or synMuv Eri suppressor genes can impact the spatiotemporal expression of GFP tagged-ergo-1 in animals. The GFP tagged-ERGO-1 fusion protein was strongly expressed in the seam cells during the L3 stage and later in the vulval epithelial cells in the mid-L4 and young adult stages (S3A–S3F Fig). We targeted the synMuv B genes lin-9 and lin-13 (S3G–S3N Fig), and the synMuv suppressor gene isw-1 by RNAi (S3O–S3R Fig) and found that the ergo-1 expression pattern was unchanged. Thus ERGO-1 is unlikely to be a direct or indirect target of synMuv B protein regulation.
The expression pattern and subcellular localization of lin-15B and lin-35 synMuv B proteins
A detailed description of the subcellular expression/localization patterns for any synMuv B gene product has not been previously reported. We investigated the expression patterns and subcellular localization of the LIN-15B and LIN-35 proteins using LIN-15B::EGFP and LIN-35::EGFP protein fusions, both generated by Mihail Sarov via a recombineering pipeline [66]. In these fusion protein reporter strains, the synMuv B gene is tagged by a protein fusion to EGFP at its C terminus in the context of roughly 40 kb of genomic DNA cloned on a fosmid, and then subsequently biolistically transformed at a relatively low copy number [66]. These strains are likely to recapitulate the endogenous expression/localization patterns of their tagged proteins and given their low copy number representation, they may not be subject to the transgene silencing typical of high-copy number transgenes that is commonly associated with Eri-genotypes. These synMuv B GFP fusion proteins were previously used to map the binding sites of the synMuv B proteins on the promoters of genes genome wide [67]. We detected robust LIN-15B::EGFP protein localization in both the hypodermal cells and intestinal cells during the mid-L4 stage and later animals. Intriguingly, LIN-15B::EGFP is localized subnuclearly, forming punctate EGFP foci in the hypodermal and intestinal nuclei. These foci may represent genomic binding sites for client genes of the LIN-15B THAP domain transcription factor, or even more interestingly, given that the dREAM complex mediates genome amplification of chorion genes in Drosophila, particular chromosomal regions that are amplified and continue to bind to synMuv B proteins [36]. To quantitate the abundance of LIN-15B::EGFP, we quantitated GFP in particular hypodermal cells and intestinal cells from the anterior, middle, and posterior (S4 Fig). We measured LIN-15B::EGFP fluorescence intensity in these cells using Image J and the number of LIN-15B::EGFP subnuclear foci was manually scored.
These subnuclear foci of LIN-15B do not change under RNAi targeting genes such mut-16, rde-1, or rde-4, which when deactivated impair RNA interference (S5 Fig). The subnuclear foci of LIN-15B also remain unperturbed under RNAi that targets genes known to enhance RNAi such as eri-6, lin-9, lin-13, lin-35, lin-37, and lin-52 or genes that are known to suppress the enhanced response to RNAi of synMuv B mutants such as isw-1 and mes-4 (S5 Fig). Crossing lin-35(n745) null mutant into the LIN-15B::EGFP reporter strain did not change LIN-15B localization or the average number of subnuclear foci (S6A–S6D Fig). Taken together, these findings suggest that the LIN-15B localization is not coupled to its role to activate the antiviral RNAi response.
Similar observations of the lin-35/retinoblastoma expression pattern and protein localization using a lin-35::EGFP translational fusion gene revealed that LIN-35::EGFP is strongly expressed in all intestinal cells, where it localizes to the nucleus as well as to the nucleolus (S7A and S7B Fig). Each intestinal cell showed 1 or 2 LIN-35::EGFP nucleolar inclusions as well as uniform localization across the nucleus (S7A and S7B Fig). There were no obvious changes to LIN-35 localization or expression level in strains induced to have an enhanced or defective RNAi by inactivation of eri or mutator/rde genes (S7C–S7E Fig).
Discussion
The potency of an injected 1 kb dsRNA for C. elegans RNA interference, discovered by Fire and Mello almost 30 years ago, was the tip of a very large RNAi iceberg. In fact, the complexity of the endogenous small RNA world immediately emerged from the genetic analysis of RNAi: the first RNAi-defective mutants identified included a large set of known mutator genes that had already emerged from searches for mutations that enhance transposition or reversion of transposons; defects in RNA interference was the molecular basis of many mutator mutants that release transposons from RNAi control [1,4,5,7,68,69]. Mutants that disable RNAi in turn activate transposable elements, integrated retroviruses in the C. elegans genome, the remnants of past successful retroviral infections that are normally silenced by RNAi, which now become active if key RNAi components are defective to then insert or revert from an insertion in any gene [68]. Thus, RNAi is not just a response to in vitro produced dsRNAs which are injected, their biological clients are natural endogenous dsRNAs that produce siRNAs which the same genetic pathways mediate their activities.
Similarly, the C. elegans enhanced RNAi mutants have highlighted a similar physiological regulation of antiviral defense or transposon mobilizations; enhanced RNAi or Eri mutants have emerged from genetic screens at about the same frequency as RNAi defective mutations (Rde, Mut, etc.). Each of the mutant classes, RNAi defective (Rde) and enhanced RNAi (Eri), points to these phenotypes as physiological states, with the mutants frozen in a state of either no RNAi defense or higher than normal RNAi defense. Conversely, Saccharomyces cerevisiae jettisoned its RNAi pathway to allow its permanent infection by the killer RNA virus [70].
The synMuv B genes emerged from large genetic and RNAi screens for enhanced RNAi mutants or gene inactivations [7,11], but were discovered 20 years earlier in unrelated comprehensive genetic analysis of receptor tyrosine kinase signaling during C. elegans vulval development [71,72]. Our analysis of the mRNA expression profiles of synMuv B null mutants showed a significant overlap between genes up-regulated in the synMuv B mutants and those induced upon viral infection by the Orsay RNA virus [41]. Notably, the 100× or more induction of genes in synMuv B mutants that are also strongly induced by Orsay virus, for example, members of the PALS gene family, suggests a potential link between synMuv B mutations and the activation of a coordinated genetic response akin to that observed during viral infections. The dramatic up-regulation of Orsay virus response genes in synMuv B mutants, even in the absence of viral infection, suggested that the down-regulation of synMuv B gene activity could mimic a physiological state of viral resistance, enhanced RNAi, that is normally induced in an actual viral infection, to activate antiviral defense mechanisms, including RNAi. The robust induction of pals-5p::GFP expression in synMuv B mutants, even in the absence of actual viral infection, suggests that these mutants constitutively activate their antiviral defense pathways.
Because the C. elegans synMuv B mutants lin-35 and lin-15B activate highly congruent gene expression responses to those of an Orsay RNA virus infection, we tested whether an actual Orsay infection down-regulates synMuv B activity: whether an Orsay virus infection can induce a Muv phenotype in a synMuv A mutant, just as a synMuv B; synMuv A double mutant shows the Muv phenotype. We found that Orsay viral infection of a single synMuv A mutant that never shows the Muv phenotype causes a reproducible 10% penetrant Muv phenotype, but Orsay viral infection of wild-type animals does not cause a Muv phenotype (shown in Fig 5A and 5B). And in support of the model that decreased synMuv B gene activity is part of the normal C. elegans antiviral response, we found that synMuv B mutants have 50 to 130× lower viral RNA levels during an Orsay virus infection than wild type (shown in Fig 5C). Thus, down-regulation of synMuv B activity to enhance the RNAi is a key component in the defense response to viral infection. Recently, Tecle and colleagues also observed a reduction in viral RNA levels in a different lin-15B mutant than what was used in our study [73]. Thus, synMuv B gene activity is integral to defense in an actual viral infection and that viral infection induces a physiological state akin to the enhanced RNAi observed in multiple synMuv B mutants.
Why is the penetrance of the Muv phenotype not higher than 10% during a viral infection of a synMuv A mutant, when null mutations in synMuv B in the background of a null synMuv A mutation can cause 100% penetrant Muv phenotype? The most likely explanation is that a viral infection lowers synMuv B activity but not to zero as in a synMuv B null mutant, that the synMuv B activity decrease during a viral infection is most similar to a non-null synMuv B mutant. In addition, the Muv phenotype of a synMuv A; synMuv B double mutant is caused by the loss of the synMuv B gene specifically in the hypodermis; the loss of synMuv B in the hypodermis is specifically required for the Muv phenotype of a synMuv A; synMuv B double mutant [22]. The loss of synMuv B and synMuv A in the hypodermis causes more than 100× increased expression of the LIN-3 ligand which is secreted to the adjacent affect vulval cells to cause the Muv phenotype in those vulval cells [15]. Because Orsay virus infection induces pals-5p::GFP in the intestine, the primary tissue where the virus infection is thought to occur via its ingestion [42], it is possible that synMuv B down-regulation in the hypodermis during a viral infection of the intestine is far less than a synMuv B null mutant, thus the lower penetrance of the Muv phenotype.
We also explored the link between gene inactivations that suppress the enhanced RNAi in synMuv B mutants and their activation of antiviral response genes. RNAi inhibition of known suppressors of the enhanced RNAi phenotype of synMuv B genes, such as isw-1 and mes-4, caused a significant reduction in pals-5p::GFP induction by a synMuv B mutation, indicating a potential interplay between RNAi and antiviral defense pathways regulated by synMuv B genes. Interestingly, our findings suggest that the Eri phenotype induced by synMuv B mutations may engage distinct pathways from other known Eri mutants, such as mutations in the RNA dependent RNA polymerase rrf-3 or the eri-1 exonuclease, highlighting the complexity of antiviral defense mechanisms in C. elegans. The observed suppression of pals-5p::GFP expression upon isw-1 and mes-4 knockdown in synMuv B mutants under non-infection conditions underscores the role of these genes in modulating the antiviral response. However, inactivation of isw-1 or mes-4 did not suppress the induction of pals-5p::GFP during an actual viral infection, suggesting that synMuv B induction of the pals-5p::GFP response, may be one of several pathways that regulates the expression of pals-5 during an actual viral infection.
C. elegans RNAi pathways surveil thousands of genomic loci in wild-type C. elegans under non-viral infection conditions to produce hundreds to thousands of siRNAs per million reads. Most of this massive cloud of siRNAs against their own genes are missing in animals with defects in RNAi, most dramatically in the Mutator mutants [30]. In contrast, null mutations in the enhanced RNAi mutant eri-6/7, encoding an RNA helicase, disrupts the production of a far more circumscribed set of siRNAs that silence approximately 100 integrated retroviral elements [10–13]. This Eri mutant may be enhanced for RNAi because the desilenced retroviral elements trigger ER stress, probably due to the replication of viral sequences in the ER, and the enhanced RNAi of an actual or falsely sensed infection [10]. Our survey of the small RNA landscape in synMuv B Eri mutants showed a 5-fold increase in 10% CSR-class 22G siRNAs, 5-fold decrease in 50% of ERGO-class 26G siRNAs, 7% of WAGO-class 22G siRNAs, 4% of piRNAs, and 14% microRNAs in synMuv B mutants. The partial loss of ERGO-26Gs and piRNAs in synMuv B mutants is distinct from the previously characterized eri mutants [12,58]. One model for the molecular basis of enhanced RNAi mutants is that there is a limiting component to RNA interference; for example, the silencing of transposons uses some limiting factor for RNAi such that if the silencing of retrotransposons is disabled, that limiting factor is released for the dsRNA-programmed silencing of feeding RNAi [64]. However, the differences in the small RNA landscape of the synMuv B mutants suggest a distinct mechanism may underlie the enhanced RNAi of synMuv B mutants: the targets of ERGO-class 26Gs, piRNAs and WAGO-class 22Gs include pseudogenes, foreign elements and transposons, loss of these small RNAs in synmuvB mutants may cause the misexpression of these genes and may explain the constitutively active anti-viral readiness of the synmuvB mutants. It is possible that the de-repression of integrated viruses, the activation of transposons, in the synMuv B mutants in fact causes a viral activation to in turn induced stronger RNAi. The induction of enhanced RNAi during a viral infection makes excellent biological sense.
The synMuv B genes encode homologues of the dREAM complex, which are conserved across animals and plants, including the LIN-35/retinoblastoma (Rb) tumor suppressor, and the Rb complex components LIN-53 /RbAp48, LIN-37/MIP40, LIN-54/MIP120, DPL-1 /DP, LIN-9/MIP130 [16,19–21]. The synMuv A genes lin-8, lin-56, and lin-15A encode widely expressed C. elegans nuclear proteins with no clear homologues outside of nematodes [17,18], and in the case of lin-8, there exists more than a dozen paralogues in C. elegans and many dozen in C. japonicum, located in clusters of duplicated paralogues [46]. The nuclear localization of the synMuv A proteins is consistent with their strong genetic interaction with the nuclearly localized synMuv B dREAM complex proteins, but nothing in the anonymous synMuv A protein sequences suggests a mechanism of action.
While the analysis of the synMuv A and synMuv B mutants in specification of the ventral precursor cells has been extensive and beautiful, how it connects to the enhanced RNAi of the synMuv B mutants is not instantly obvious from the beautiful molecular genetic analysis of LIN-3 signaling [15,16,20,21,23]. The molecular basis of how the DNA-binding proteins and associated proteins of the synMuv B pathway actually intersect with the PIWI proteins and RdRPs that are central to RNAi has not been characterized. Our investigations suggested that RNAi suppression of the Eri-phenotype of synMuv B mutants by isw-1 or mes-4 RNAi do not affect the localization of LIN-15B or LIN-35 (see S5 and S7 Figs). This suggests that a molecular mechanism distinct from the localization or abundance of the synMuv B proteins regulates the intensity of the RNAi pathway.
However, extensive analysis of the many synMuv B orthologues in Drosophila has revealed their activity in endoreduplication of particular client genes, and the detailed analysis of C. elegans synMuv B genes in the vulval cell signaling pathway has revealed an analogous intersection with endoreduplication. The Drosophila dREAM complex binds specifically at replication origins that flank the Drosophila chorion genes to control their local (about 50 kb) endoreduplication to nearly 100× the gene dosage of the normally diploid Drosophila genome [36]. This enables the regulated production of the abundant eggshell proteins; mutations in the dREAM complex cause a humpty dumpty phenotype, fragile eggs due to insufficient production of chorion proteins [38,74]. The dREAM complex proteins bind directly to these chorion gene replication origins and are necessary for the localized amplification of just those genes; in the Drosophila retinoblastoma mutant, unregulated endoreduplication occurs [37–39].
This intersection of the dREAM complex with endoreduplication is also a hallmark of the synMuv B gene function in LIN-3 growth factor signaling from the polyploid C. elegans hypodermis to the adjacent vulval cells that show the Muv phenotype in the synMuv B mutants. The polyploid hypodermal cells hyp7 cell is the focus of synMuv B gene function in C. elegans for the regulation of LIN-3 production [15]. The C. elegans hypoderm is normally endoreduplicated to 8C (3 full genome duplications) or more at adult stages [75].
Analysis of other shared C. elegans synMuv B mutant phenotypes also suggests that they also function in another polyploid tissue, the intestine, to control the production of P-granules which are approximately 1 micron ribonucleoprotein granules intimately connected to RNAi [30]. The P-granules are normally germline-specific multi-protein and RNAs that assemble into the originally described LLPS particles of the single blastomere that is the precursor of the C. elegans germline [28]. Loss of function mutations in multiple C. elegans synMuv B genes cause misexpression of these normally germline-specific P-granules in somatic tissues, most dramatically in the polyploid intestine and hypodermis [7,29]. The adult C. elegans intestine is normally endoreduplicated from diploid at hatching to 32C (5 inferred full genome duplications with the DAPI-light microscope DNA quantitation), with one duplication at each larval stage but no chromosome condensation or mitosis [25]. The hyp7 hypoderm where ectopic P granules are also observed is 8C. The P granules, as macromolecular complexes that are visible at light microscope magnification, are filled with abundant proteins and RNAs, which like chorion proteins, represent candidate genes for localized endoreduplication [29,31–34,76].
It may be significant that nearly all of the 50 genes that are strongly up-regulated in synMuv B mutants and during viral infection are clustered in the genome (see S1 Fig and the S5 Data). For example, the pals genes as well as the lin-8-like genes (lido-genes) localize to genomic clusters with several pals or lido genes within cluster. The clusters are also very plastic in Caenorhabditis evolution, varying down to just 3 lido genes in C. remaniae and up to 58 lido genes in C. japonicum. These clusters of poorly conserved pals and lido genes may be under dREAM complex control of their local amplification, which may serve to increase the gene copy number for example in the intestine.
That RNAi is highly regulated is not surprising: RNAi has evolved by selection for potent antiviral defense during almost a billion years of eukaryotic infection by viruses. But it is surprising that the dREAM complex is so central to RNAi. And the mammalian-conserved synMuv B pathway reveals new pathways to activate production or response to small interfering RNAs and may be targeted with drugs to activate human antiviral responses or responses to RNAi-based pharmaceuticals.
Materials and methods
Nematode strains used
Strains made available through the CAENORHABDITIS GENETICS CENTER (CGC): N2 (wild-type), lin-35(n745), lin-52(n771), lin-9(n112), lin-15B(n744), lin-15AB(n765), eri-6/7(mg441), eri-1(mg366), NL2099 [rrf-3(pk1426], ERT54 [jyIs8 [pals-5p::GFP; myo-2p::mCherry], mgIs30 [rol-6(su1006)::lin-14 3′-UTR, col-10::lacZ, lim-6::gfp]), OP184[wgIs184 [lin-15B::TY1::EGFP::3xFLAG + unc-119(+)], OP763[wgIs763 [lin-35::TY1::EGFP::3xFLAG + unc-119(+)], lin-8(n2731), qIs56(lag-2::gfp, unc-119(þ), eri-1(mg366), eri-6(mg379), rrf-3(pk1426), mut-16(pk710).
More details regarding the gene mutations featured here can be found at http://www.wormbase.org.
Strains generated in our lab for this study: lin-35(n745) glp-4(bn2), glp-4(bn2); lin-15B(n744), glp-4(bn2), lin-35(n745); jyIs8, lin-52(n771); jyIs8, lin-15B(W485*), lin-15B(W485*); jj2284, lin-15B(W485*);FAS207, eri-1(mg366); jyIs8, eri-6(mg379); jyIs8, rrf-3(pk1426); jyIs8, lin-9(n112); qIs56, lin-35(n745); qIs56.
Strains sent by others*: FAS207, unc-119(ed3) III; zuIs261[ges-1(5′ UTR)::npp-9::mCherry::BLRP::3xFLAG::npp-9(3’UTR); unc-119(+)]; zuIs236[his-72(5’ UTR)::BIRA::GFP::his-72(3’ UTR); unc-119(+)].
JJ2284 unc-119(ed3) III; zuIs261[ges-1(5’ UTR)::npp-9::mCherry::BLRP::3xFLAG::npp-9(3’ UTR); unc-119(+). *These strains were kindly provided by Steven Henikoff and Florian Steiner.
Generating the strain lin-15B(W485*): In brief, gravid animals were injected with a CRISPR/Cas9 mix containing 30 pmol S. pyrogenes Cas9 (IDT), 90 pmol tracrRNA (IDT), 95 pmol crRNA oHG6, 2.2 mg repair template oHG8, and 80 ng of PRF4::rol-6 (su1006) plasmid. The F1 progeny that exhibited the rol phenotype as well as their siblings were singled and genotyped for the edit using primers oHG9 and oHG10, followed by a restriction digest using AseI (New England Biolabs).
Oligos used
oHG6: AGAATGAGAATTCGAATCAT
oHG8: GAGCGGCCCGAAGAGAATGAGAATTCGAATCATTAATGGGGAATGCGTCAGACGGATATGGAATTCTTGGC
oHG9: ACAAAAACTTTTATCAAGATATTTCGATCCTG
oHG10: ACAAAAACTTTTATCAAGATATTTCGATCCTG
Counting intestinal nuclei: Animals infected with Orsay virus were collected in M9, crushed and filtered through a 0.22 μm filter; 50 μl of the resulting lysate was used to infect animals. Equal volume of M9 was used for the mock infection. Four L4 animals were infected with mock or Orsay virus lysate. Animals were allowed to propagate for 2 generations. When the animals were running out of food, the animals were chunked to fresh plates pre-infected with the Orsay virus lysate. F2 adults were picked into sodium azide on an agarose pad and imaged on the Zeiss Axio Imager Z.1 within an hour. The number of mCherry expressing intestinal nuclei was scored using the 40× objective.
RT-qPCR for Orsay RNA1: Growth conditions: Ten 60 mm plates of mut-16(pk710) animals infected with Orsay virus and the OP50 lawn that they were grown on were collected in approximately 8.5 ml of M9. Worms were spun down and the supernatant was moved to a fresh tube. The worms were moved to a 1.5 ml microcentrifuge tube and crushed using a pestle. The worm debris was spun down and the resulting clear lysate was added to the previously collected supernatant. The pooled supernatant was filtered through a 0.22 μm filter and 350 μl of the resulting filtrate was used to infect approximately 20 fresh 60 mm plates each. Equal volume of M9 was used for mock infection. Adults of the relevant genotypes were bleached using sodium hypochlorite treatment and embryos were allowed to hatch on the infected lawns. Approximately 1,200 animals were collected in Trizol 3 days after plating as adults (N2 and eri-1) or late L4s (mut-16, lin-15b, lin-35).
RNA extraction and RT-qPCR: Samples were freeze–thawed thrice using liquid nitrogen. After the addition of 1-bromo-3-chloropropane, the RNA in the aqueous phase was precipitated by incubating with isopropanol for 2 h at −30°C. Samples were centrifuged at 21,000 × g for 30 min at 4°C to pellet the RNA. The pellet was washed thrice in 70% ethanol, dried and resuspended in water; 500 ng of RNA was used to make cDNA with BIO-RAD iScript cDNA Synthesis Kit (cat# 1708890) according to manufacturer’s instructions, and 2 μl of the resulting cDNA was used for Quantitative PCR with BIO-RAD iQ SYBR Green Supermix (cat# 170–8880). The primers used for amplifying eft-2 and Orsay RNA1 are listed at the end of this section. The cycle numbers for Orsay RNA1 were normalized to the respective cycle numbers for eft-2. Two biological replicates with 2 technical replicates were done. The values were all normalized to the average of the 4 values for the wild-type N2 infected samples. Two-tailed Student’s unpaired t test was done to evaluate p-values.
Oligos used
oHG17: ACGCTCGTGATGAGTTCAAG [qPCR FP for eft-2]
oHG18: ATTTGGTCCAGTTCCGTCTG [qPCR RP for eft-2]
oHG53: ACCTCACAACTGCCATCTACA [qPCR FP for Orsay RNA1 [48]]
oHG50: GACGCTTCCAAGATTGGTATTGGT [qPCR RP for Orsay RNA1 [48]]
siRNA Sequencing: Growth conditions
Eggs were hatched and arrested as synchronous L1 larvae and then plated on E. coli OP50 and grown at either 20°C for 78 to 96 h (glp-4+) or 25°C for 72 to 74 h (glp-4-). Samples were collected for RNA isolation when most animals had reached adult stage, which varied between strains. There was some stage asynchrony within the SynMuv mutant strains because of differences in development timing.
RNA isolation: RNA was isolated from whole animals using Trizol and chloroform extraction according to the manufacturer’s recommendations but with the addition of a second chloroform extraction step (Life Technologies, cat# 15596018).
Small RNA sequencing: Total RNA was treated with RppH to reduce small RNA 5′ triphosphates to monophosphates following the manufacturer’s recommendations (New England Biolabs, cat# M0356S) [77]; 16 to 30-nt small RNAs were size selected on a 17% polyacrylamide/urea gel and purified using the ZR small-RNA PAGE Recovery Kit (Zymo Research, cat# R1070). Small RNA sequencing libraries were prepared using the NEBNext Multiplex Small RNA Library Prep Set for Illumina following the manufacturer’s recommendations but with the 3′ ligation step changed to 16°C for 18 h to improve capture of methylated small RNAs (New England Biolabs, cat# E7300S). PCR products corresponding in size to adapter-ligated small RNAs were size selected on a 10% polyacrylamide non-denaturing gel, electrophoretically transferred to DE81 chromatography paper, eluted at 70°C for 20 min in the presence of 1 M NaCl, and precipitated at −80°C overnight in the presence of 13 μg/ml glycogen and 67% EtOH. Small RNA read processing, including adapter trimming, quality filtering, mapping, and counting, was done with tinyRNA v1.5 using the default configuration [55]. Reference sequences and annotations were based on the C. elegans WS279 release [78].
GEO accession numbers and genotype information for the siRNAseq data
Project accession number: GSE283983
Individual sample accession numbers: GSM8675299 (N2 rep 1), GSM8675300 (N2 rep 2), GSM8675301(lin-15b rep 1), GSM8675302 (lin-15b rep 2),GSM8675303 (lin-35 rep 1), GSM8675304 (lin-35 rep 2), GSM8675305 (lin-52 rep 1), GSM8675306 (lin-52 rep 2), GSM8675307 (lin-9 rep 1), GSM8675308 (lin-9 rep 2), GSM8675309 (glp-4 rep 1), GSM8675310 (glp-4 rep 2), GSM8675311 (lin-15b glp-4 rep 1), GSM8675312 (lin-15b glp-4 rep 2), GSM8675313 (lin-35 glp-4 rep 1), GSM8675314 (lin-35 glp-4 rep 2).
Generating the top 100 up and down list of siRNA targets
- DESeq2 analysis using R was performed on 3 sample groups at once (i.e., glp-4; lin-15B and lin-35 glp-4 and glp-4) for the 0 mm (mismatch) set.
- Pairwise comparisons (i.e., lin-15B; glp-4 and glp-4 OR lin-35; glp-4 and glp-4) yielded log2FoldChange and adjusted p-values (padj). To identify the siRNAs with significant changes up in the synMuv B mutants, the list of siRNA targets was filtered as follows:
- ○ Greater than or equal to 5-fold change (i.e., log2FoldChange = log2(5) = 2.32);
- ○ Less than or equal to adjusted p-value of 0.05; and
- ○ Greater than or equal to 10 counts of normalized reads in the synMuv B; glp-4 strains.
- To identify the siRNAs with significant changes down in the synMuv B mutants, the list of siRNA targets was filtered as follows:
- ○ Less than or equal to 0.2-fold change (i.e., log2FoldChange = log2(0.2) = −2.32);
- ○ Less than or equal to adjusted p-value of 0.05; and
- ○ Greater than or equal to 10 counts of normalized reads in the glp-4 strains.
- Each of the lists of siRNA targets were arranged in the order of greatest fold change (i.e., greatest log2FoldChange for siRNA targets that are the most up-regulated and lowest log2FoldChange for siRNA targets that are the most down-regulated).
mRNA sequencing analysis
Fastq files were downloaded from GEO (GEO Accession numbers GSE62833 and GSE155190). The STAR aligner [79] was used to map sequencing reads to transcriptome in the ce10 reference genome. Read counts for individual genes were produced using the count function in HTSeq [80], followed by the estimation of expression values and detection of differentially expressed transcripts using EdgeR [81], which included only the genes with count per million reads (CPM) > 1 for 2 or more samples. Differentially expressed genes were defined by the cutoffs of > log2 fold change and false discovery rate (FDR) S1 Fig was generated using venn and matplotlib in Python 3.
GEO accession numbers and genotype information
GSM4697084 (N2 starved L1-rep1), GSM4697085 (N2 starved L1-rep2), GSM4697089 [lin-35(n745) starved L1-rep1)], GSM4697090 (lin-35(n745) starved L1-rep2), GSM4697102 (lin-15B(n744) starved L1-rep1), GSM4697105 (lin-15B(n744) starved L1-rep2), GSM1534086 (lin-35[JA1507(n745) L3-rep1), GSM1534087(lin-35[JA1507(n745) L3-rep2).
Supporting information
S1 Fig. Gene expression analysis reveals antiviral defense genes that are up-regulated in synMuv B mutants.
Most genes (80/91) up-regulated during Orsay virus infection of wild-type animals [41] are also up-regulated in synMuvB mutants [82,83]. Venn diagram representing overlap between the published data sets is on the left; 46 genes that are up-regulated in all datasets is listed on the right. All genes analyzed here were up-regulated by at least 5-fold compared to the respective control samples. The S5 Data shows that most of the up-regulated genes this analysis revealed are clustered in the genome, indicative of local regulation of gene expression or perhaps even gene endoreduplication by synMuv B genes.
https://doi.org/10.1371/journal.pbio.3002748.s001
(TIF)
S2 Fig. Ectopic lag-2p::GFP expression in the intestinal cells upon virus-infection remains unaltered upon removal of isw-1 activity by RNAi.
(A) Fluorescent micrographs showing 2 lag-2p::GFP expressing animals that are infected with the Orsay virus raised on E. coli that express double stranded RNA against either empty vector (control) or the synMuv suppressor gene isw-1. Expression of lag-2p::GFP within the distal tip cells is indicated by yellow arrow heads and the ectopic fluorescence of lag-2p::GFP in the intestine is also indicated. (B) Quantification of the fluorescence intensity of lag-2p::GFP within the intestine of animals that are infected with the Orsay virus raised on E. coli that express double stranded RNA against either the L4440 empty vector (labeled as control) or isw-1 is shown. A schematic of the general orientation of the worm is shown above the image panels. Scale bar is indicated. The raw microscopy images shown in this figure have been deposited in Zenodo and are accessible at DOI: 10.5281/zenodo.14289232.
https://doi.org/10.1371/journal.pbio.3002748.s002
(TIF)
S3 Fig. Spatiotemporal expression pattern of ERGO-1 remains unperturbed by the loss of function of synMuv B genes.
(A–F) Brightfield and GFP channel micrographs depicting the temporal expression pattern of ergo-1::GFP from a transgenic strain that expresses a full-length ERGO-1 PIWI protein fused at its C terminus to GFP. (G–R) ergo-1::GFP expression after synMuv B (lin-9 or lin-13), or Muv suppressor (isw-1) RNAi. Developmental stage is labeled. Scale bar is indicated. The raw microscopy images shown in this figure have been deposited in Zenodo and are accessible at DOI: 10.5281/zenodo.14289232.
https://doi.org/10.1371/journal.pbio.3002748.s003
(TIF)
S4 Fig. LIN-15B C-terminal protein fusion::EGFP is localized to subnuclear foci in C. elegans polyploid hypodermal and intestinal cells.
(A–E) Brightfield and EGFP micrographs of LIN-15B localization in representative hypodermal (panels A and B), intestinal nuclei (panels C–E). Abbreviations for the different intestinal cells that are boxed are provided below the image panels. Scale bar is indicated. The raw microscopy images shown in this figure have been deposited in Zenodo and are accessible at DOI: 10.5281/zenodo.14289232.
https://doi.org/10.1371/journal.pbio.3002748.s004
(TIF)
S5 Fig. Quantification of LIN-15B::EGFP fluorescence intensity and number of subnuclear foci.
(A, B) Quantification of EGFP fluorescence and number of subnuclear LIN-15B foci in 3 representative hypodermal cells for synMuv B (lin-9, lin-13, lin-35, lin-37 and lin-52), Eri associated-gene (eri-6), synMuv suppressors (mes-4, isw-1), RNAi defective genes (rde-1, rde-4, mut-16) under RNAi conditions. (C, D) Quantification of EGFP fluorescence and number of subnuclear LIN-15B foci within 6 representative intestinal cells for synMuv B (lin-9, lin-13, lin-35, lin-37 and lin-52), Eri associated-gene (eri-6), synMuv suppressors (mes-4, isw-1), RNAi defective genes (rde-1, rde-4, mut-16) under RNAi conditions. In the graphs shown in C and D the abbreviations AD refers to anterior dorsal cell, AV refers to anterior ventral cell, MA refers to midgut anterior cell and MP refers to midgut posterior cell, PD refers to posterior gut dorsal cell and PV refers to posterior gut ventral cell. The numerical data underlying the graphs shown in this figure can be found in the Supporting information data table named S1 Raw Data.
https://doi.org/10.1371/journal.pbio.3002748.s005
(TIF)
S6 Fig. Quantification of LIN-15B::EGFP fusion protein fluorescence intensity and number of subnuclear foci in lin-35(n745) null mutants.
(A, B) EGFP fluorescence and number of subnuclear LIN-15B foci within 3 representative hypodermal cells under RNAi conditions targeting L4440 vector (control), RNAi defective genes (rde-1, mut-16), and Muv suppressor (isw-1 and mes-4). (C, D) Intensity of EGFP fluorescence and number of subnuclear LIN-15B foci within 6 representative intestinal cells where the abbreviations AD refers to anterior dorsal cell, AV refers to anterior ventral cell, MA refers to midgut anterior cell and MP refers to midgut posterior cell, PD refers to posterior gut dorsal cell, and PV refers to posterior gut ventral cell. RNAi was performed targeting L4440 vector (control), RNAi defective genes (rde-1, mut-16), and Muv suppressor genes (isw-1 and mes-4). These experiments were carried out in a lin-35(n745) null background. The numerical data underlying the graphs shown in this figure can be found in the Supporting information data table named S1 Raw Data.
https://doi.org/10.1371/journal.pbio.3002748.s006
(TIF)
S7 Fig. LIN-35::EGFP localization in the nucleolus of C. elegans intestinal cells.
(A, B) Fluorescent micrographs of LIN-35::EGFP fusion protein localization in the intestinal nucleoli of wild-type animals. Magnification used to visualize the cells is indicated. The image shown in panel B are the same cells that are boxed in panel A, visualized under a higher magnification. (C) Fluorescent micrographs of LIN-35::EGFP localization within the intestinal cells (boxed in panel A), of the F1 progeny of worms raised on E. coli expressing double stranded RNA against either empty vector (l4440) or various synMuv B genes (lin-9, lin-13, lin-15b, lin-37, lin-52, lin-54, lin-61, tam-1, hpl-1, and dpl-1) or a synMuv A gene (lin-8) is shown. Arrow heads indicate the presence of a nucleolar inclusion of LIN-35::EGFP. (D) Quantification of the number of LIN-35::EGFP nucleolar inclusions that are seen in animals raised on E. coli expressing double stranded RNA against either empty vector (l4440) or various synMuv B genes (lin-9, lin-13, lin-15b, lin-37, lin-52, lin-54, lin-61, tam-1, hpl-1, and dpl-1) or a synMuv A gene (lin-8) is shown. (E) Quantification of the number of LIN-35::EGFP nucleolar inclusions that are seen in animals raised on E. coli expressing double stranded RNA against either empty vector (l4440) or various genes that are critical for the worms ability to perform RNAi. Scale bar is indicated. The raw microscopy images shown in this figure have been deposited in Zenodo and are accessible at DOI: 10.5281/zenodo.14289232.
https://doi.org/10.1371/journal.pbio.3002748.s007
(TIF)
S8 Fig. lin-15b(-) animals exhibit altered intestinal nuclear morphology.
Top panel shows wild-type JJ2284 animals under no virus and Orsay virus-infected conditions. Lower panel shows a second independent line of lin-15b(W485*); JJ2284 animals under no virus and Orsay virus-infected conditions. White arrows indicate elongated intestinal nuclei that are elongated. Scale bar is indicated. The raw microscopy images shown in this figure have been deposited in Zenodo and are accessible at DOI: 10.5281/zenodo.14289232.
https://doi.org/10.1371/journal.pbio.3002748.s008
(TIF)
S9 Fig. Comparisons of normalized reads of indicated classes of small RNAs in lin-35(n745); glp-4(bn2) with glp-4(bn2).
Dark gray data points represent small RNAs that are differentially expressed by 5-fold and adjusted p-value lin-35(n745); glp-4(bn2) (upper) and glp-4(bn2) (lower). The numerical data underlying the graphs shown in this figure can be found in the Supporting information data table named S1 Raw Data.
https://doi.org/10.1371/journal.pbio.3002748.s009
(TIF)
S10 Fig. Comparisons of normalized reads of indicated classes of small RNAs in lin-15b(n744); glp-4(bn2) with glp-4(bn2).
Dark gray data points represent small RNAs that are differentially expressed by 5-fold and adjusted p-value lin-15b(n744); glp-4(bn2) (upper) and glp-4(bn2) (lower). The numerical data underlying the graphs shown in this figure can be found in the Supporting information data table named S1 Raw Data.
https://doi.org/10.1371/journal.pbio.3002748.s010
(TIF)
S11 Fig. Comparisons of normalized reads of indicated classes of small RNAs in lin-35(n745) with wild type.
Dark gray data points represent small RNAs that are differentially expressed by 5-fold and adjusted p-value lin-35(n745) (upper) and wild type (lower). The numerical data underlying the graphs shown in this figure can be found in the Supporting information data table named S1 Raw Data.
https://doi.org/10.1371/journal.pbio.3002748.s011
(TIF)
S12 Fig. Comparisons of normalized reads of indicated classes of small RNAs in lin-15b(n744) with wild type.
Dark gray data points represent small RNAs that are differentially expressed by 5-fold and adjusted p-value lin-15b(n744) (upper) and wild type (lower). The numerical data underlying the graphs shown in this figure can be found in the Supporting information data table named S1 Raw Data.
https://doi.org/10.1371/journal.pbio.3002748.s012
(TIF)
S13 Fig. Comparisons of normalized reads of indicated classes of small RNAs in lin-9(n112) with wild type.
B Dark gray data points represent small RNAs that are differentially expressed by 5-fold and adjusted p-value lin-9(n112) (upper) and wild type (lower). The numerical data underlying the graphs shown in this figure can be found in the Supporting information data table named S1 Raw Data.
https://doi.org/10.1371/journal.pbio.3002748.s013
(TIF)
S14 Fig. Comparisons of normalized reads of indicated classes of small RNAs in lin-52(n771) with wild type.
Dark gray data points represent small RNAs that are differentially expressed by 5-fold and adjusted p-value lin-52(n771)(upper) and wild type (lower). The numerical data underlying the graphs shown in this figure can be found in the Supporting information data table named S1 Raw Data.
https://doi.org/10.1371/journal.pbio.3002748.s014
(TIF)
S1 Table. Top 50 up-regulated genes in lin-15B(n744), mutant in starved L1 animals.
The 50 most highly up-regulated genes in lin-15(n744) mutants during the L1 stage. The log2 Fold-change and the corresponding false discovery rate (FDR), value for each gene that is listed is shown. The gene expression data was derived from our analysis of mRNA seq data sets [GSM4697084- N2—Rep 1/L1, GSM4697085- N2—Rep 2/L1 against GSM4697102- lin-15b(n744)- rep1/L1, GSM4697105- lin-15b(n744)- rep2/L1] that were obtained from the NCBI Geo collection.
https://doi.org/10.1371/journal.pbio.3002748.s015
(XLSX)
S2 Table. Top 50 up-regulated genes in lin-35(n745), mutant in starved L1 animals.
The 50 most highly up-regulated genes in lin-35(n745) mutants during the L1 stage. The log2 Fold-change and the corresponding false discovery rate (FDR), value for each gene that is listed is shown. The gene expression data was derived from our analysis of mRNA seq data sets GSM4697089- lin-35[JA1507(n745) rep1/L1, GSM4697090- lin-35[JA1507(n745) rep1/L1] that were obtained from the NCBI Geo collection.
https://doi.org/10.1371/journal.pbio.3002748.s016
(XLSX)
S3 Table. Top 50 up-regulated Genes in lin-35(n745), mutant at the L3 Stage.
The top 50 most highly up-regulated genes in lin-35(n745) mutants during the L3 stage. The log2 Fold-change and the corresponding false discovery rate (FDR), value for each gene that is listed is shown. [GSM1534084- N2-rep1 L3, GSM1534085- N2-rep2 compared against GSM1534086- lin-35[JA1507(n745) rep1 L3 and GSM1534087- lin-35[JA1507(n745) rep1 L3] that were obtained from the NCBI Geo collection.
https://doi.org/10.1371/journal.pbio.3002748.s017
(XLSX)
S1 Data. List of genes to which differentially regulated small RNAs map in lin-35(n745); glp-4(bn2) and lin-15b(n744); glp-4(bn2) in comparison with glp-4(bn2).
The siRNAs that are either Up or Down by a minimum factor of a 5-fold difference in the lin-15(n744) and lin-35(n745) mutant backgrounds are presented. See the labeled tabs for the genotype information.
https://doi.org/10.1371/journal.pbio.3002748.s018
(XLSX)
S2 Data. Gene expression changes in lin-35(n745) starved L1 animals.
Shown here are the lists of significant gene expression changes via mRNA seq that are observed in the lin-35(n745) synMuv B mutant animals at the L1 stage. The data shown here was obtained from the NCBI geo collection, GSM4697089 [lin-35(n745) starved L1-rep1)], GSM4697090 (lin-35(n745) starved L1-rep2)].
https://doi.org/10.1371/journal.pbio.3002748.s019
(XLS)
S3 Data. Gene expression changes in lin-35(n745) starved L3-stage animals.
Shown here are the lists of significant gene expression changes via mRNA seq that are observed in the lin-35(n745) synMuv B mutant animals at the L3-stage. The data shown here was obtained from the NCBI geo collection, GSM1534086 (lin-35[JA1507(n745) L3-rep1), GSM1534087(lin-35[JA1507(n745) L3-rep2)].
https://doi.org/10.1371/journal.pbio.3002748.s020
(XLS)
S4 Data. Gene expression changes in lin-15(n744) starved L1-stage animals.
Shown here are the lists of significant gene expression changes via mRNA seq that are observed in the lin-15(n744) synMuv B mutant animals at the L1-stage. The data shown here was obtained from the NCBI geo collection, GSM4697102 (lin-15B(n744) starved L1-rep1), GSM4697105 (lin-15B(n744) starved L1-rep2)].
https://doi.org/10.1371/journal.pbio.3002748.s021
(XLS)
References
- 1.
Ketting RF, Cochella L. Concepts and functions of small RNA pathways in C. elegans. Curr Top Dev Biol. 2021;144:45–89. pmid:33992161 - 2.
Timmons L, Fire A. Specific interference by ingested dsRNA. Nature. 1998;395(6705):854. pmid:9804418 - 3.
Vastenhouw NL, Fischer SE, Robert VJ, Thijssen KL, Fraser AG, Kamath RS, et al. A genome-wide screen identifies 27 genes involved in transposon silencing in C. elegans. Curr Biol. 2003;13(15):1311–6. pmid:12906791 - 4.
Ketting RF, Haverkamp TH, van Luenen HG, Plasterk RH. Mut-7 of C. elegans, required for transposon silencing and RNA interference, is a homolog of Werner syndrome helicase and RNaseD. Cell. 1999;99(2):133–41. pmid:10535732 - 5.
Simmer F, Tijsterman M, Parrish S, Koushika SP, Nonet ML, Fire A, et al. Loss of the putative RNA-directed RNA polymerase RRF-3 makes C. elegans hypersensitive to RNAi. Curr Biol. 2002;12(15):1317–9. pmid:12176360 - 6.
Wu X, Shi Z, Cui M, Han M, Ruvkun G. Repression of germline RNAi pathways in somatic cells by retinoblastoma pathway chromatin complexes. PLoS Genet. 2012;8(3):e1002542. pmid:22412383 - 7.
Wang D, Kennedy S, Conte D Jr., Kim JK, Gabel HW, Kamath RS , et al. Somatic misexpression of germline P granules and enhanced RNA interference in retinoblastoma pathway mutants. Nature. 2005;436(7050):593–7. pmid:16049496 - 8.
Lipsick JS. synMuv verite—Myb comes into focus. Genes Dev. 2004;18(23):2837–44. pmid:15574590 - 9.
Gabel HW, Ruvkun G. The exonuclease ERI-1 has a conserved dual role in 5.8S rRNA processing and RNAi. Nat Struct Mol Biol. 2008;15(5):531–3. pmid:18438419 - 10.
Fischer SEJ, Ruvkun G. Caenorhabditis elegans ADAR editing and the ERI-6/7/MOV10 RNAi pathway silence endogenous viral elements and LTR retrotransposons. Proc Natl Acad Sci U S A. 2020;117(11):5987–96. pmid:32123111 - 11.
Fischer SE, Pan Q, Breen PC, Qi Y, Shi Z, Zhang C, et al. Multiple small RNA pathways regulate the silencing of repeated and foreign genes in C. elegans. Genes Dev. 2013;27(24):2678–95. pmid:24352423 - 12.
Fischer SE, Montgomery TA, Zhang C, Fahlgren N, Breen PC, Hwang A, et al. The ERI-6/7 helicase acts at the first stage of an siRNA amplification pathway that targets recent gene duplications. PLoS Genet. 2011;7(11):e1002369. pmid:22102828 - 13.
Fischer SE, Butler MD, Pan Q, Ruvkun G. Trans-splicing in C. elegans generates the negative RNAi regulator ERI-6/7. Nature. 2008;455(7212):491–6. pmid:18784652 - 14.
Cui M, Kim EB, Han M. Diverse chromatin remodeling genes antagonize the Rb-involved SynMuv pathways in C. elegans. PLoS Genet. 2006;2(5):e74. pmid:16710447 - 15.
Saffer AM, Kim DH, van Oudenaarden A, Horvitz HR. The Caenorhabditis elegans synthetic multivulva genes prevent ras pathway activation by tightly repressing global ectopic expression of lin-3 EGF. PLoS Genet. 2011;7(12):e1002418. pmid:22242000 - 16.
Clark SG, Lu X, Horvitz HR. The Caenorhabditis elegans locus lin-15, a negative regulator of a tyrosine kinase signaling pathway, encodes two different proteins. Genetics. 1994;137(4):987–97. pmid:7982579 - 17.
Davison EM, Harrison MM, Walhout AJ, Vidal M, Horvitz HR. lin-8, which antagonizes Caenorhabditis elegans Ras-mediated vulval induction, encodes a novel nuclear protein that interacts with the LIN-35 Rb protein. Genetics. 2005;171(3):1017–31. pmid:16020796 - 18.
Davison EM, Saffer AM, Huang LS, DeModena J, Sternberg PW, Horvitz HR. The LIN-15A and LIN-56 transcriptional regulators interact to negatively regulate EGF/Ras signaling in Caenorhabditis elegans vulval cell-fate determination. Genetics. 2011;187(3):803–15. pmid:21196525 - 19.
Lu X, Horvitz HR. lin-35 and lin-53, two genes that antagonize a C. elegans Ras pathway, encode proteins similar to Rb and its binding protein RbAp48. Cell. 1998;95(7):981–91. pmid:9875852 - 20.
Harrison MM, Lu X, Horvitz HR. LIN-61, one of two Caenorhabditis elegans malignant-brain-tumor-repeat-containing proteins, acts with the DRM and NuRD-like protein complexes in vulval development but not in certain other biological processes. Genetics. 2007;176(1):255–71. pmid:17409073 - 21.
Harrison MM, Ceol CJ, Lu X, Horvitz HR. Some C. elegans class B synthetic multivulva proteins encode a conserved LIN-35 Rb-containing complex distinct from a NuRD-like complex. Proc Natl Acad Sci U S A. 2006;103(45):16782–7. pmid:17075059 - 22.
Myers TR, Greenwald I. lin-35 Rb acts in the major hypodermis to oppose ras-mediated vulval induction in C. elegans. Dev Cell. 2005;8(1):117–23. pmid:15621535 - 23.
Cui M, Chen J, Myers TR, Hwang BJ, Sternberg PW, Greenwald I, et al. SynMuv genes redundantly inhibit lin-3/EGF expression to prevent inappropriate vulval induction in C. elegans. Dev Cell. 2006;10(5):667–72. pmid:16678779 - 24.
Hedgecock EM, Herman RK. The ncl-1 gene and genetic mosaics of Caenorhabditis elegans. Genetics 1995;141(3):989–1006. pmid:8582642 - 25.
Hedgecock EM, White JG. Polyploid tissues in the nematode Caenorhabditis elegans. Dev Biol. 1985;107(1):128–33. pmid:2578115 - 26.
Pitt JN, Schisa JA, Priess JR. P granules in the germ cells of Caenorhabditis elegans adults are associated with clusters of nuclear pores and contain RNA. Dev Biol. 2000;219(2):315–33. pmid:10694425 - 27.
Gu W, Shirayama M, Conte D, Jr., Vasale J, Batista PJ, Claycomb JM, et al. Distinct argonaute-mediated 22G-RNA pathways direct genome surveillance in the C. elegans germline. Mol Cell. 2009;36(2):231–44. pmid:19800275 - 28.
Brangwynne CP, Eckmann CR, Courson DS, Rybarska A, Hoege C, Gharakhani J, et al. Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science. 2009;324(5935):1729–32. pmid:19460965 - 29.
Petrella LN, Wang W, Spike CA, Rechtsteiner A, Reinke V, Strome S. synMuv B proteins antagonize germline fate in the intestine and ensure C. elegans survival. Development. 2011;138(6):1069–79. pmid:21343362 - 30.
Phillips CM, Montgomery TA, Breen PC, Ruvkun G. MUT-16 promotes formation of perinuclear mutator foci required for RNA silencing in the C. elegans germline. Genes Dev. 2012;26(13):1433–44. pmid:22713602 - 31.
Ni JZ, Kalinava N, Chen E, Huang A, Trinh T, Gu SG. A transgenerational role of the germline nuclear RNAi pathway in repressing heat stress-induced transcriptional activation in C. elegans. Epigenetics Chromatin. 2016;9:3. pmid:26779286 - 32.
Phillips CM, Montgomery BE, Breen PC, Roovers EF, Rim YS, Ohsumi TK, et al. MUT-14 and SMUT-1 DEAD box RNA helicases have overlapping roles in germline RNAi and endogenous siRNA formation. Curr Biol. 2014;24(8):839–44. pmid:24684932 - 33.
Wan G, Fields BD, Spracklin G, Shukla A, Phillips CM, Kennedy S. Spatiotemporal regulation of liquid-like condensates in epigenetic inheritance. Nature. 2018;557(7707):679–83. pmid:29769721 - 34.
Kirienko NV, Fay DS. Transcriptome profiling of the C. elegans Rb ortholog reveals diverse developmental roles. Dev Biol. 2007;305(2):674–84. pmid:17368442 - 35.
Herman RK, Hedgecock EM. Limitation of the size of the vulval primordium of Caenorhabditis elegans by lin-15 expression in surrounding hypodermis. Nature. 1990;348(6297):169–71. pmid:2234080 - 36.
Spradling AC. The organization and amplification of two chromosomal domains containing Drosophila chorion genes. Cell. 1981;27(1 Pt 2):193–201. pmid:6799210 - 37.
Bosco G, Du W, Orr-Weaver TL. DNA replication control through interaction of E2F-RB and the origin recognition complex. Nat Cell Biol. 2001;3(3):289–95. pmid:11231579 - 38.
Bandura JL, Beall EL, Bell M, Silver HR, Botchan MR, Calvi BR. humpty dumpty is required for developmental DNA amplification and cell proliferation in Drosophila. Curr Biol. 2005;15(8):755–9. pmid:15854909 - 39.
Rotelli MD, Bolling AM, Killion AW, Weinberg AJ, Dixon MJ, Calvi BR. An RNAi Screen for Genes Required for Growth of Drosophila Wing Tissue. G3 (Bethesda). 2019;9(10):3087–100. pmid:31387856 - 40.
Kennedy S, Wang D, Ruvkun G. A conserved siRNA-degrading RNase negatively regulates RNA interference in C. elegans. Nature. 2004;427(6975):645–9. pmid:14961122 - 41.
Chen K, Franz CJ, Jiang H, Jiang Y, Wang D. An evolutionarily conserved transcriptional response to viral infection in Caenorhabditis nematodes. BMC Genomics. 2017;18(1):303. pmid:28415971 - 42.
Reddy KC, Dror T, Sowa JN, Panek J, Chen K, Lim ES, et al. An Intracellular Pathogen Response Pathway Promotes Proteostasis in C. elegans. Curr Biol. 2017;27(22):3544–53 e5. - 43.
Mishra P, Ngo J, Ashkani J, Pio F. Meta-analysis suggests evidence of novel stress-related pathway components in Orsay virus—Caenorhabditis elegans viral model. Sci Rep. 2019;9(1):4399. pmid:30867481 - 44.
Lazetic V, Blanchard MJ, Bui T, Troemel ER. Multiple pals gene modules control a balance between immunity and development in Caenorhabditis elegans. PLoS Pathog. 2023;19(7):e1011120. pmid:37463170 - 45.
Leyva-Diaz E, Stefanakis N, Carrera I, Glenwinkel L, Wang G, Driscoll M, et al. Silencing of Repetitive DNA Is Controlled by a Member of an Unusual Caenorhabditis elegans Gene Family. Genetics. 2017;207(2):529–45. pmid:28801529 - 46.
Rahe DP, Hobert O. Restriction of Cellular Plasticity of Differentiated Cells Mediated by Chromatin Modifiers, Transcription Factors and Protein Kinases. G3 (Bethesda). 2019;9(7):2287–302. pmid:31088904 - 47.
Ishizu H, Siomi H, Siomi MC. Biology of PIWI-interacting RNAs: new insights into biogenesis and function inside and outside of germlines. Genes Dev. 2012;26(21):2361–73. pmid:23124062 - 48.
Felix MA, Ashe A, Piffaretti J, Wu G, Nuez I, Belicard T, et al. Natural and experimental infection of Caenorhabditis nematodes by novel viruses related to nodaviruses. PLoS Biol. 2011;9(1):e1000586. pmid:21283608 - 49.
Henderson ST, Gao D, Christensen S, Kimble J. Functional domains of LAG-2, a putative signaling ligand for LIN-12 and GLP-1 receptors in Caenorhabditis elegans. Mol Biol Cell. 1997;8(9):1751–62. pmid:9307971 - 50.
Kramer JM, Johnson JJ. Analysis of mutations in the sqt-1 and rol-6 collagen genes of Caenorhabditis elegans. Genetics. 1993;135(4):1035–45. pmid:8307321 - 51.
Ashe A, Sarkies P, Le Pen J, Tanguy M, Miska EA. Antiviral RNA Interference against Orsay Virus Is neither Systemic nor Transgenerational in Caenorhabditis elegans. J Virol. 2015;89(23):12035–46. pmid:26401037 - 52.
Ashe A, Belicard T, Le Pen J, Sarkies P, Frezal L, Lehrbach NJ, et al. A deletion polymorphism in the Caenorhabditis elegans RIG-I homolog disables viral RNA dicing and antiviral immunity. Elife. 2013;2:e00994. pmid:24137537 - 53.
Collins J, Saari B, Anderson P. Activation of a transposable element in the germ line but not the soma of Caenorhabditis elegans. Nature. 1987;328(6132):726–8. pmid:3039378 - 54.
Vasale JJ, Gu W, Thivierge C, Batista PJ, Claycomb JM, Youngman EM, et al. Sequential rounds of RNA-dependent RNA transcription drive endogenous small-RNA biogenesis in the ERGO-1/Argonaute pathway. Proc Natl Acad Sci U S A. 2010;107(8):3582–7. pmid:20133583 - 55.
Tate AJ, Brown KC, Montgomery TA. tiny-count: a counting tool for hierarchical classification and quantification of small RNA-seq reads with single-nucleotide precision. Bioinform Adv. 2023;3(1):vbad065. pmid:37288323 - 56.
Reinke V, Gil IS, Ward S, Kazmer K. Genome-wide germline-enriched and sex-biased expression profiles in Caenorhabditis elegans. Development. 2004;131(2):311–23. pmid:14668411 - 57.
Gent JI, Schvarzstein M, Villeneuve AM, Gu SG, Jantsch V, Fire AZ, et al. A Caenorhabditis elegans RNA-directed RNA polymerase in sperm development and endogenous RNA interference. Genetics. 2009;183(4):1297–314. pmid:19805814 - 58.
Han T, Manoharan AP, Harkins TT, Bouffard P, Fitzpatrick C, Chu DS, et al. 26G endo-siRNAs regulate spermatogenic and zygotic gene expression in Caenorhabditis elegans. Proc Natl Acad Sci U S A. 2009;106(44):18674–9. pmid:19846761 - 59.
Conine CC, Batista PJ, Gu W, Claycomb JM, Chaves DA, Shirayama M, et al. Argonautes ALG-3 and ALG-4 are required for spermatogenesis-specific 26G-RNAs and thermotolerant sperm in Caenorhabditis elegans. Proc Natl Acad Sci U S A. 2010;107(8):3588–93. pmid:20133686 - 60.
Claycomb JM, Batista PJ, Pang KM, Gu W, Vasale JJ, van Wolfswinkel JC, et al. The Argonaute CSR-1 and its 22G-RNA cofactors are required for holocentric chromosome segregation. Cell. 2009;139(1):123–34. pmid:19804758 - 61.
Gerson-Gurwitz A, Wang S, Sathe S, Green R, Yeo GW, Oegema K, et al. A Small RNA-Catalytic Argonaute Pathway Tunes Germline Transcript Levels to Ensure Embryonic Divisions. Cell. 2016;165(2):396–409. pmid:27020753 - 62.
Chaves DA, Dai H, Li L, Moresco JJ, Oh ME, Conte D Jr, et al. The RNA phosphatase PIR-1 regulates endogenous small RNA pathways in C. elegans. Mol Cell. 2021;81(3):546–57 e5. - 63.
Billi AC, Fischer SE, Kim JK. Endogenous RNAi pathways in C. elegans. WormBook. 2014:1–49. - 64.
Yigit E, Batista PJ, Bei Y, Pang KM, Chen CC, Tolia NH, et al. Analysis of the C. elegans Argonaute family reveals that distinct Argonautes act sequentially during RNAi. Cell. 2006;127(4):747–57. pmid:17110334 - 65.
Seroussi U, Lugowski A, Wadi L, Lao RX, Willis AR, Zhao W, et al. A comprehensive survey of C. elegans argonaute proteins reveals organism-wide gene regulatory networks and functions. Elife. 2023;12. pmid:36790166 - 66.
Sarov M, Schneider S, Pozniakovski A, Roguev A, Ernst S, Zhang Y, et al. A recombineering pipeline for functional genomics applied to Caenorhabditis elegans. Nat Methods. 2006;3(10):839–44. pmid:16990816 - 67.
Gerstein MB, Lu ZJ, Van Nostrand EL, Cheng C, Arshinoff BI, Liu T, et al. Integrative analysis of the Caenorhabditis elegans genome by the modENCODE project. Science. 2010;330(6012):1775–87. pmid:21177976 - 68.
Robert VJ, Sijen T, van Wolfswinkel J, Plasterk RH. Chromatin and RNAi factors protect the C. elegans germline against repetitive sequences. Genes Dev. 2005;19(7):782–7. pmid:15774721 - 69.
Ketting RF, Plasterk RH. A genetic link between co-suppression and RNA interference in C. elegans. Nature. 2000;404(6775):296–8. pmid:10749214 - 70.
Drinnenberg IA, Fink GR, Bartel DP. Compatibility with killer explains the rise of RNAi-deficient fungi. Science. 2011;333(6049):1592. pmid:21921191 - 71.
Ceol CJ, Stegmeier F, Harrison MM, Horvitz HR. Identification and classification of genes that act antagonistically to let-60 Ras signaling in Caenorhabditis elegans vulval development. Genetics. 2006;173(2):709–26. pmid:16624904 - 72.
Fay DS, Yochem J. The SynMuv genes of Caenorhabditis elegans in vulval development and beyond. Dev Biol. 2007;306(1):1–9. pmid:17434473 - 73.
Tecle E, Li S, Blanchard MJ, Bui T, Chhan CB, Underwood RS, et al. Conserved chromatin regulators control the transcriptional immune response to intracellular pathogens in Caenorhabditis elegans. bioRxiv. 2024.10.03.616425. - 74.
Lewis PW, Beall EL, Fleischer TC, Georlette D, Link AJ, Botchan MR. Identification of a Drosophila Myb-E2F2/RBF transcriptional repressor complex. Genes Dev. 2004;18(23):2929–40. pmid:15545624 - 75.
Flemming AJ, Shen ZZ, Cunha A, Emmons SW, Leroi AM. Somatic polyploidization and cellular proliferation drive body size evolution in nematodes. Proc Natl Acad Sci U S A. 2000;97(10):5285–90. pmid:10805788 - 76.
Kelly AL, Senter-Zapata MJ, Campbell AC, Lust HE, Theriault ME, Andralojc KM, et al. A Forward Genetic Screen for Suppressors of Somatic P Granules in Caenorhabditis elegans. G3 (Bethesda). 2015;5(10):2209–15. pmid:26100681 - 77.
Almeida MV, de Jesus Domingues AM, Lukas H, Mendez-Lago M, Ketting RF. RppH can faithfully replace TAP to allow cloning of 5’-triphosphate carrying small RNAs. MethodsX. 2019;6:265–72. pmid:30788220 - 78.
Davis P, Zarowiecki M, Arnaboldi V, Becerra A, Cain S, Chan J, et al. WormBase in 2022-data, processes, and tools for analyzing Caenorhabditis elegans. Genetics. 2022;220(4). pmid:35134929 - 79.
Dobin A, Davis CA, Schlesinger F, Drenkow J, Zaleski C, Jha S, et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics. 2013;29(1):15–21. pmid:23104886 - 80.
Putri GH, Anders S, Pyl PT, Pimanda JE, Zanini F. Analysing high-throughput sequencing data in Python with HTSeq 2.0. Bioinformatics. 2022;38(10):2943–5. pmid:35561197 - 81.
Robinson MD, McCarthy DJ, Smyth GK. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics. 2010;26(1):139–40. pmid:19910308 - 82.
Gal C, Carelli FN, Appert A, Cerrato C, Huang N, Dong Y, et al. DREAM represses distinct targets by cooperating with different THAP domain proteins. Cell Rep. 2021;37(3):109835. pmid:34686342 - 83.
Latorre I, Chesney MA, Garrigues JM, Stempor P, Appert A, Francesconi M, et al. The DREAM complex promotes gene body H2A.Z for target repression. Genes Dev. 2015;29(5):495–500. pmid:25737279
ADVERTISEMENT:
Hai, sobat pengemar slots Pernah mendengar istilah “slot demo”? jika tidak, siap-siap jatuh cinta dengan program ini. slot demo adalah mesin slots yang sering kasih win. Yup, slot-slot ini bisa disebut adalah andalannya tuk membawa come back hasil. but, cemana sih caranya nemuin slot gacor yang tepat? Santai Bro, kita bahas tenang saja di sini
Gaming terpopuler saat ini hanya satu di Indonesia yaitu yang menyediakan imbal hasil terbaik
SEGERA hanya di :
Informasi mengenai KING SLOT, Segera Daftar Bersama king selot terbaik dan terpercaya no satu di Indonesia. Boleh mendaftar melalui sini king slot serta memberikan hasil kembali yang paling tinggi saat sekarang ini hanyalah KING SLOT atau Raja slot paling gacor, gilak dan gaco saat sekarang di Indonesia melalui program return tinggi di kingselot serta pg king slot
slot demo gacor
slot demo gacor permainan paling top dan garansi imbal balik hasil besar bersama kdwapp.com
akun demo slot gacor
akun demo slot gacor permainan paling top dan garansi imbal balik hasil besar bersama kdwapp.com
akun slot demo gacor
akun slot demo gacor permainan paling top dan garansi imbal balik hasil besar bersama kdwapp.com
akun demo slot pragmatic
akun demo slot pragmatic permainan paling top dan garansi imbal balik hasil besar bersama kdwapp.com
akun slot demo pragmatic
akun slot demo pragmatic permainan paling top dan garansi imbal balik hasil besar bersama kdwapp.com
akun slot demo
akun slot demo permainan paling top dan garansi imbal balik hasil besar bersama kdwapp.com
akun demo slot
akun demo slot permainan paling top dan garansi imbal balik hasil besar bersama kdwapp.com
slot demo gacor
slot demo gacor permainan paling top dan garansi imbal balik hasil besar bersama jebswagstore.com
akun demo slot gacor
akun demo slot gacor permainan paling top dan garansi imbal balik hasil besar bersama jebswagstore.com
akun slot demo gacor
akun slot demo gacor permainan paling top dan garansi imbal balik hasil besar bersama jebswagstore.com
akun demo slot pragmatic
akun demo slot pragmatic permainan paling top dan garansi imbal balik hasil besar bersama jebswagstore.com
akun slot demo pragmatic
akun slot demo pragmatic permainan paling top dan garansi imbal balik hasil besar bersama jebswagstore.com
akun slot demo
akun slot demo permainan paling top dan garansi imbal balik hasil besar bersama jebswagstore.com
akun demo slot
akun demo slot permainan paling top dan garansi imbal balik hasil besar bersama jebswagstore.com
slot demo gacor
slot demo gacor permainan paling top dan garansi imbal balik hasil besar bersama demoslotgacor.pro
akun demo slot gacor
akun demo slot gacor permainan paling top dan garansi imbal balik hasil besar bersama demoslotgacor.pro
akun slot demo gacor
akun slot demo gacor permainan paling top dan garansi imbal balik hasil besar bersama demoslotgacor.pro
akun demo slot pragmatic
akun demo slot pragmatic permainan paling top dan garansi imbal balik hasil besar bersama demoslotgacor.pro
akun slot demo pragmatic
akun slot demo pragmatic permainan paling top dan garansi imbal balik hasil besar bersama demoslotgacor.pro
akun slot demo
akun slot demo permainan paling top dan garansi imbal balik hasil besar bersama demoslotgacor.pro
akun demo slot
akun demo slot permainan paling top dan garansi imbal balik hasil besar bersama demoslotgacor.pro
slot demo gacor
slot demo gacor permainan paling top dan garansi imbal balik hasil besar bersama situsslotterbaru.net
akun demo slot gacor
akun demo slot gacor permainan paling top dan garansi imbal balik hasil besar bersama situsslotterbaru.net
akun slot demo gacor
akun slot demo gacor permainan paling top dan garansi imbal balik hasil besar bersama situsslotterbaru.net
akun demo slot pragmatic
akun demo slot pragmatic permainan paling top dan garansi imbal balik hasil besar bersama situsslotterbaru.net
akun slot demo pragmatic
akun slot demo pragmatic permainan paling top dan garansi imbal balik hasil besar bersama situsslotterbaru.net
akun slot demo
akun slot demo permainan paling top dan garansi imbal balik hasil besar bersama situsslotterbaru.net
akun demo slot
akun demo slot permainan paling top dan garansi imbal balik hasil besar bersama situsslotterbaru.net
situs slot terbaru
situs slot terbaru permainan paling top dan garansi imbal balik hasil besar bersama situsslotterbaru.net
slot terbaru
slot terbaru permainan paling top dan garansi imbal balik hasil besar bersama situsslotterbaru.net
suara88 permainan paling top dan garansi imbal balik hasil besar bersama suara88.biz
sumo7777 permainan paling top dan garansi imbal balik hasil besar bersama sumo7777.com
supermoney888 permainan paling top dan garansi imbal balik hasil besar bersama supermoney888.biz
teratai88 permainan paling top dan garansi imbal balik hasil besar bersama teratai88.biz
thor88 permainan paling top dan garansi imbal balik hasil besar bersama thor88.biz
togelhk88 permainan paling top dan garansi imbal balik hasil besar bersama togelhk88.net
topjitu88 permainan paling top dan garansi imbal balik hasil besar bersama topjitu88.net
totosloto88 permainan paling top dan garansi imbal balik hasil besar bersama totosloto88.com
trisula888 permainan paling top dan garansi imbal balik hasil besar bersama trisula888.biz
udangbet88 permainan paling top dan garansi imbal balik hasil besar bersama udangbet88.net
via88 permainan paling top dan garansi imbal balik hasil besar bersama via88.biz
virusjp88 permainan paling top dan garansi imbal balik hasil besar bersama virusjp88.net
warga888 permainan paling top dan garansi imbal balik hasil besar bersama warga888.biz
waw88 permainan paling top dan garansi imbal balik hasil besar bersama waw88.biz
winjitu88 permainan paling top dan garansi imbal balik hasil besar bersama winjitu88.net
wisdom88 permainan paling top dan garansi imbal balik hasil besar bersama wisdom88.biz
wnitogel88 permainan paling top dan garansi imbal balik hasil besar bersama wnitogel88.com
yoyo888 permainan paling top dan garansi imbal balik hasil besar bersama yoyo888.biz
validtoto88 permainan paling top dan garansi imbal balik hasil besar bersama validtoto88.com
sule999 permainan paling top dan garansi imbal balik hasil besar bersama sule999.com
sule88 permainan paling top dan garansi imbal balik hasil besar bersama sule88.org
ss888bet permainan paling top dan garansi imbal balik hasil besar bersama ss888bet.com
sia77 permainan paling top dan garansi imbal balik hasil besar bersama sia77.info
seluang88 permainan paling top dan garansi imbal balik hasil besar bersama seluang88.com
satu88 permainan paling top dan garansi imbal balik hasil besar bersama satu88.biz
satu777 permainan paling top dan garansi imbal balik hasil besar bersama satu777.asia
rp88 permainan paling top dan garansi imbal balik hasil besar bersama rp88.biz
rp88 permainan paling top dan garansi imbal balik hasil besar bersama rp88.asia
rp88 permainan paling top dan garansi imbal balik hasil besar bersama rp77.live
qiuqiu88 permainan paling top dan garansi imbal balik hasil besar bersama qiuqiu88.biz
pt88 permainan paling top dan garansi imbal balik hasil besar bersama pt88.org
pt77 permainan paling top dan garansi imbal balik hasil besar bersama pt77.info
produk88 permainan paling top dan garansi imbal balik hasil besar bersama produk88.asia
mt88 permainan paling top dan garansi imbal balik hasil besar bersama mt88.org
mt77 permainan paling top dan garansi imbal balik hasil besar bersama mt77.biz
menang66 permainan paling top dan garansi imbal balik hasil besar bersama menang66.biz
latobet888 permainan paling top dan garansi imbal balik hasil besar bersama latobet888.org
kedai96 permainan paling top dan garansi imbal balik hasil besar bersama kedai96.org
kedai188 permainan paling top dan garansi imbal balik hasil besar bersama kedai188.biz
ids88 permainan paling top dan garansi imbal balik hasil besar bersama ids88.biz
hp88 permainan paling top dan garansi imbal balik hasil besar bersama hp88.org
hp77 permainan paling top dan garansi imbal balik hasil besar bersama hp77.org
gm88 permainan paling top dan garansi imbal balik hasil besar bersama gm88.asia
gm77 permainan paling top dan garansi imbal balik hasil besar bersama gm77.net
final888 permainan paling top dan garansi imbal balik hasil besar bersama final888.org
duit88 permainan paling top dan garansi imbal balik hasil besar bersama duit88.asia
duit168 permainan paling top dan garansi imbal balik hasil besar bersama duit168.biz
divisi88 permainan paling top dan garansi imbal balik hasil besar bersama divisi88.org
dewi500 permainan paling top dan garansi imbal balik hasil besar bersama dewi500.biz
devil88 permainan paling top dan garansi imbal balik hasil besar bersama devil88.info
cuputoto88 permainan paling top dan garansi imbal balik hasil besar bersama cuputoto88.com
cukongbet88 permainan paling top dan garansi imbal balik hasil besar bersama cukongbet88.asia
bom888 permainan paling top dan garansi imbal balik hasil besar bersama bom888.biz
bintaro888 permainan paling top dan garansi imbal balik hasil besar bersama bintaro888.info
askasino88 permainan paling top dan garansi imbal balik hasil besar bersama askasino88.org
999aset permainan paling top dan garansi imbal balik hasil besar bersama 999aset.com
afb77 permainan paling top dan garansi imbal balik hasil besar bersama afb77.biz
aset99 permainan paling top dan garansi imbal balik hasil besar bersama aset99.biz
bendera77 permainan paling top dan garansi imbal balik hasil besar bersama bendera77.biz
bendera888 permainan paling top dan garansi imbal balik hasil besar bersama bendera888.com
coco88 permainan paling top dan garansi imbal balik hasil besar bersama coco88.org
cuma77 permainan paling top dan garansi imbal balik hasil besar bersama cuma77.biz
cuma88 permainan paling top dan garansi imbal balik hasil besar bersama cuma88.org
dwv88 permainan paling top dan garansi imbal balik hasil besar bersama dwv88.org
fafajp88 permainan paling top dan garansi imbal balik hasil besar bersama fafajp88.com
gemar88 permainan paling top dan garansi imbal balik hasil besar bersama gemar88.biz
gocap88 permainan paling top dan garansi imbal balik hasil besar bersama gocap88.info
gocaptoto permainan paling top dan garansi imbal balik hasil besar bersama gocaptoto.asia
hakabet88 permainan paling top dan garansi imbal balik hasil besar bersama hakabet88.com
hwtoto88 permainan paling top dan garansi imbal balik hasil besar bersama hwtoto88.org
ina77 permainan paling top dan garansi imbal balik hasil besar bersama ina77.biz
ina88 permainan paling top dan garansi imbal balik hasil besar bersama ina88.info
jingga8888 permainan paling top dan garansi imbal balik hasil besar bersama jingga8888.com
juragan777 permainan paling top dan garansi imbal balik hasil besar bersama juragan777.asia
kastil77 permainan paling top dan garansi imbal balik hasil besar bersama kastil77.info
kebo888 permainan paling top dan garansi imbal balik hasil besar bersama kebo888.biz
kkwin77 permainan paling top dan garansi imbal balik hasil besar bersama kkwin77.com
kokoslot88 permainan paling top dan garansi imbal balik hasil besar bersama kokoslot88.asia
luckydf88 permainan paling top dan garansi imbal balik hasil besar bersama luckydf88.org
microstar888 permainan paling top dan garansi imbal balik hasil besar bersama microstar888.biz
monperatoto88 permainan paling top dan garansi imbal balik hasil besar bersama monperatoto88.com
mpo1122 permainan paling top dan garansi imbal balik hasil besar bersama mpo1122.biz
mpo122 permainan paling top dan garansi imbal balik hasil besar bersama mpo122.biz
mpopelangi88 permainan paling top dan garansi imbal balik hasil besar bersama mpopelangi88.com
pamanslot88 permainan paling top dan garansi imbal balik hasil besar bersama pamanslot88.biz
panel88 permainan paling top dan garansi imbal balik hasil besar bersama panel88.org
paragon77 permainan paling top dan garansi imbal balik hasil besar bersama paragon77.biz
paragon888 permainan paling top dan garansi imbal balik hasil besar bersama paragon888.info
pion77 permainan paling top dan garansi imbal balik hasil besar bersama pion77.biz
prada88 permainan paling top dan garansi imbal balik hasil besar bersama prada88.asia
prada888 permainan paling top dan garansi imbal balik hasil besar bersama prada888.com
qqslot88slot permainan paling top dan garansi imbal balik hasil besar bersama qqslot88slot.com
rejekibet88 permainan paling top dan garansi imbal balik hasil besar bersama rejekibet88.com
rezekibet88 permainan paling top dan garansi imbal balik hasil besar bersama rezekibet88.org
sensa77 permainan paling top dan garansi imbal balik hasil besar bersama sensa77.biz
sensa888 permainan paling top dan garansi imbal balik hasil besar bersama sensa888.biz
singajp88 permainan paling top dan garansi imbal balik hasil besar bersama singajp88.com
sr77 permainan paling top dan garansi imbal balik hasil besar bersama sr77.org
sr88 permainan paling top dan garansi imbal balik hasil besar bersama sr88.org
surya77 permainan paling top dan garansi imbal balik hasil besar bersama surya77.biz
surya88 permainan paling top dan garansi imbal balik hasil besar bersama surya88.asia
tajir77 permainan paling top dan garansi imbal balik hasil besar bersama tajir77.info
tajir88 permainan paling top dan garansi imbal balik hasil besar bersama tajir88.biz
toto122 permainan paling top dan garansi imbal balik hasil besar bersama toto122.com
toto123 permainan paling top dan garansi imbal balik hasil besar bersama toto123.biz
uangvip88 permainan paling top dan garansi imbal balik hasil besar bersama uangvip88.com
wajik77 permainan paling top dan garansi imbal balik hasil besar bersama wajik77.asia
777neko permainan paling top dan garansi imbal balik hasil besar bersama 777neko.org
88judi permainan paling top dan garansi imbal balik hasil besar bersama 88judi.net
99judi permainan paling top dan garansi imbal balik hasil besar bersama 99judi.org
abcslot88 permainan paling top dan garansi imbal balik hasil besar bersama abcslot88.asia



